






另外,成骨细胞分泌IL-6以刺激破骨细胞形成。许多血管的中膜平滑肌细胞也产生IL-6作为促炎性细胞因子。IL-6作为抗炎性肌动蛋白的作用是通过其对TNF-α和IL-1的抑制作用以及IL-1ra和IL-10的激活而介导的。
早期证据表明,在更广泛的冠状病毒大流行的背景下,IL-6可以用作严重COVID-19感染的炎性标志物,预后较差。
一、IL-6受体
IL-6通过细胞表面I型细胞因子受体复合物发出信号,该复合物由结合配体的IL-6Rα链(CD126)和信号转导成分gp130(也称为CD130)组成。CD130是多种细胞因子的常见信号转导子,包括白血病抑制因子(LIF),睫状神经营养因子,IL-11和心肌钙蛋白1,并且几乎在大多数组织中普遍表达。相反,CD126的表达仅限于某些组织。
除膜结合受体外,还从人血清和尿液中纯化了可溶形式的IL-6R(sIL-6R)。许多神经元细胞对单独的IL-6刺激无反应,但是神经元细胞的分化和存活可以通过sIL-6R的作用来介导。sIL-6R/IL-6复合物可刺激神经突长出并促进神经元的存活,因此,可能在通过髓鞘再生的神经再生中起重要作用。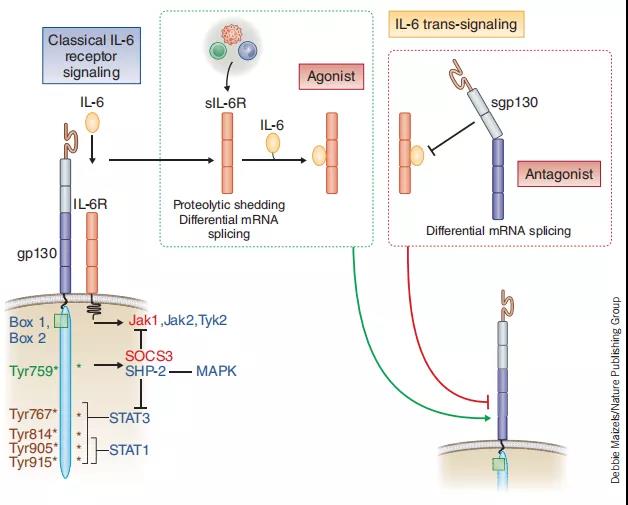
二、IL-6功能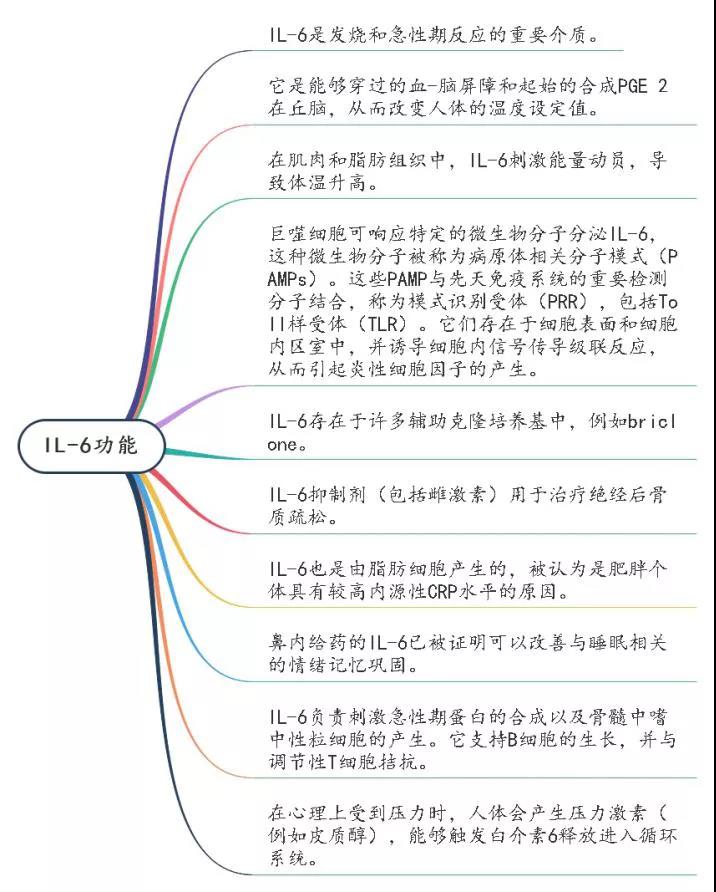
三、白细胞介素6(IL-6)(具有多效活性的细胞因子)的多效性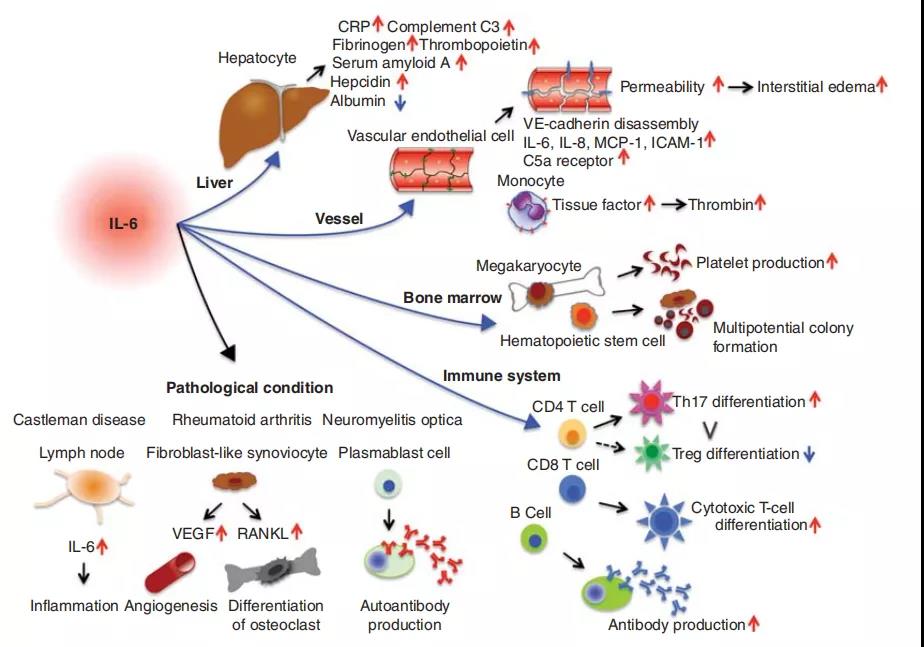
它诱导肝脏中急性期蛋白的合成,例如C反应蛋白(CRP),补体C3,纤维蛋白原,血小板生成素,血清淀粉样蛋白A和铁调素,而它抑制白蛋白的产生。IL-6通过刺激抗体产生并诱导幼稚CD4þT细胞分化为效应T细胞,在获得性免疫应答中也起重要作用。IL-6激活血管内皮细胞产生IL-6,IL-8,单核细胞趋化蛋白1(MCP-1),细胞间粘附分子(ICAM)-1和C5a受体,并诱导血管内皮钙黏着蛋白分解。另外,IL-6可以促进几种非免疫细胞的分化或增殖。由于其多效活性,IL-6持续或过量生产失调导致各种疾病的发作或发展。IL-6的过量产生在病理上与Castleman病的淋巴结肿大有关,而滑液中过量的IL-6刺激成纤维样滑膜细胞产生血管内皮生长因子(VEGF)和核因子kB(NF)受体激活剂-kB)配体(RANKL),可增强类风湿关节炎患者的血管生成和骨质疏松。IL-6支持成纤维细胞的存活,成纤维细胞在视神经脊髓炎患者中产生抗水通道蛋白4抗体。
四、不同细胞对IL-6的反应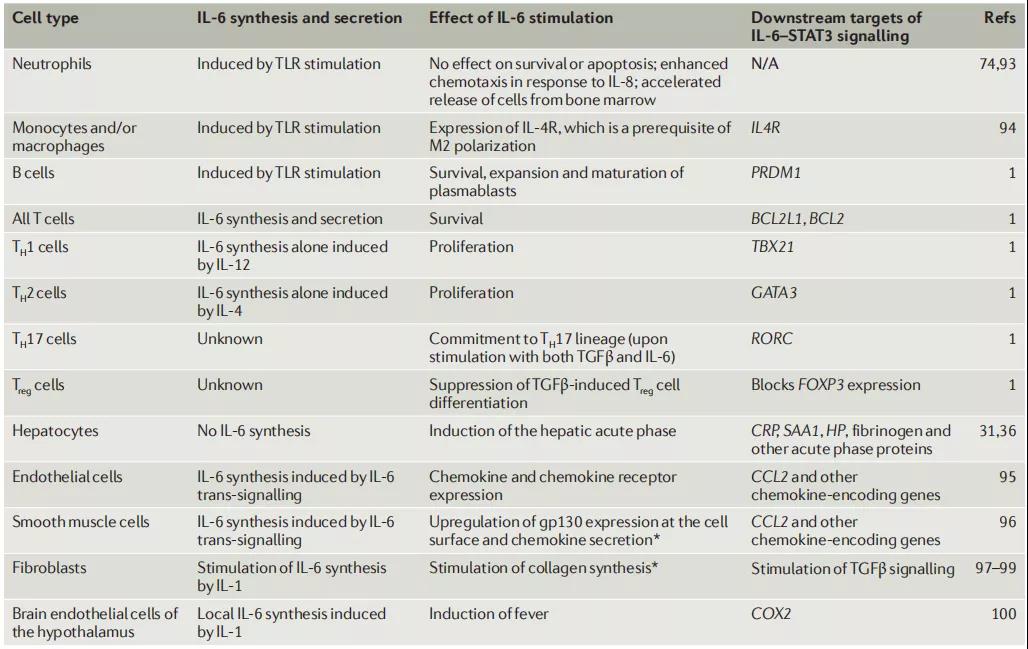
1、IL-6控制B细胞和体液免疫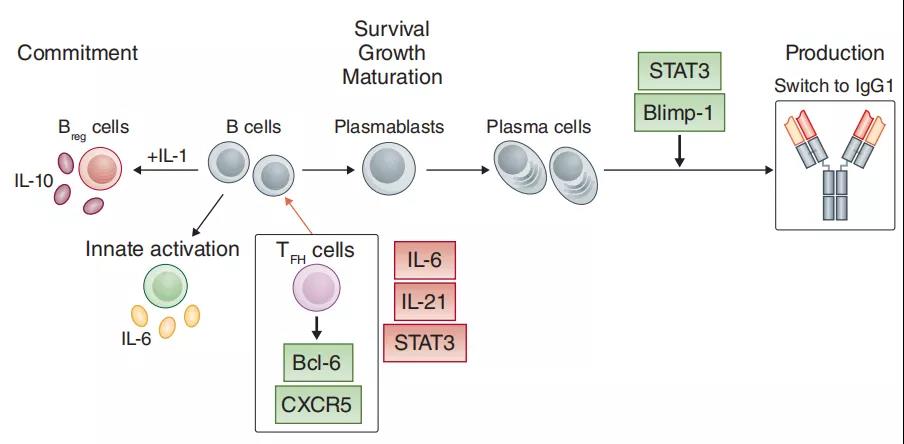
IL-6控制B细胞和成浆细胞的存活,种群扩张和成熟。例如,STAT3对转录因子Blimp-1的调节与抗体的分泌有关,并且与产生大量免疫球蛋白的长寿浆细胞相关。IL-6还控制T细胞中IL-21的表达,并且IL-6和IL-21对STAT3的激活会增强Bcl-6的表达和TFH细胞的生成。Blimp-1和Bcl-6的活性相互抵消,这种相互关系会影响淋巴细胞分化和淋巴细胞功能。通过确定的Toll样受体先天激活B细胞,也控制B细胞产生IL-6,并且IL-6与IL-1b结合可为分泌分泌IL-10-的调节性B细胞提供必要的信号。
2、IL-6对T细胞活性的影响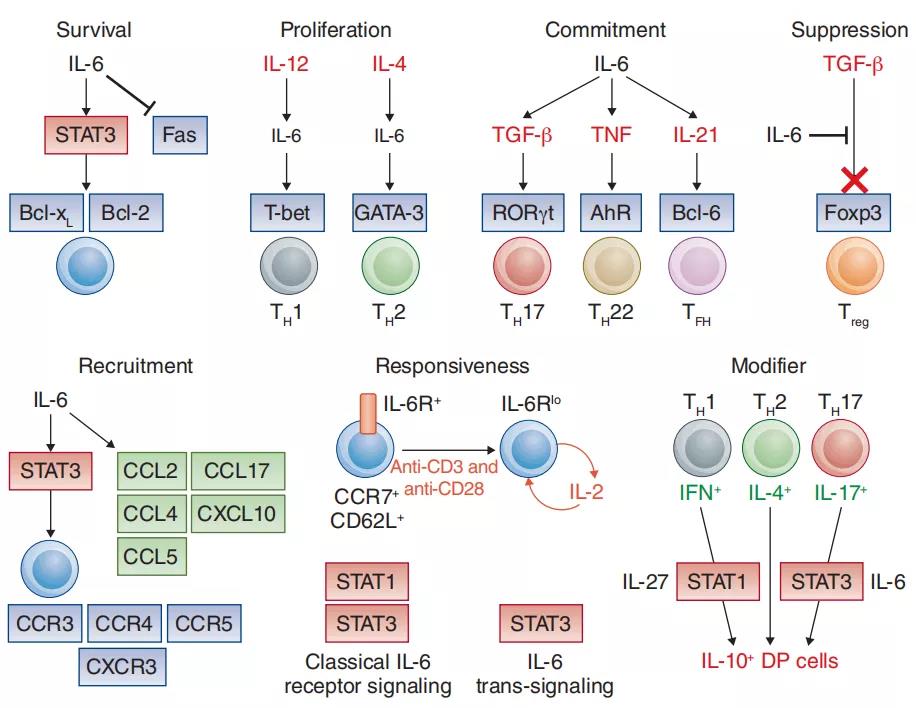
IL-6控制T细胞的增殖,存活和定向,并调节其效应细胞因子的产生。这里的例子强调了IL-6在维持T细胞应答(例如,控制TH1或TH2细胞的增殖和存活)方面的支持作用,而IL-6对于确定的效应子群体的发展至关重要。辅助性T细胞的子集TH17,TH22和TFH或其抑制作用(例如Treg细胞)。IL-6还通过控制趋化因子受体的表达来调节T细胞浸润,作用在基质组织上的IL-6反式信号传导也调节了引起T细胞募集的几种炎症趋化因子。同源IL-6R表达主要与表达CCR7和CD62L的幼稚或中央记忆T细胞相关。T细胞抗原受体的活化促进IL-6受体的脱落,并伴随着IL-6介导的STAT1活性的丧失。IL-2的存在阻止了IL-6R在T细胞表面的呈递。这表明T细胞的活化导致T细胞对IL-6的应答性的改变以及从经典的IL-6R信号转为IL-6反信号转导。IL-6还修饰了定义的T细胞群体的效应子特性。例如,IL-27和IL-6之间的信号相互作用(涉及STAT1和STAT3的激活)通过定义的效应T细胞亚群促进IL-10的分泌。类似地,IL-6还可通过诱导性Treg细胞群体获得T-bet或RORgt的表达。
3、IL-6在T细胞亚群分化中的作用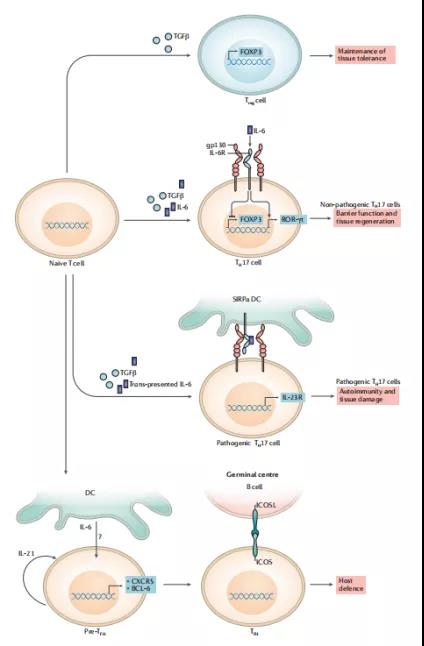
白介素-6(IL-6)是一种非冗余细胞因子,参与T辅助细胞17(TH17)细胞的分化。幼稚T细胞通过其膜结合IL-6受体(IL-6R)感知IL-6; IL-6阻止了转化生长因子-β(TGFβ)介导的FOXP3的诱导,并促进了视黄酸受体相关的孤儿核受体γ(ROR-γt)(TH17的主要转录因子)的表达细胞。然而,仅当树突状细胞(DC)将IL-6转染至T细胞时,组织破坏性TH17细胞才被引发,该树突状细胞(DC)也向T细胞提供同源(抗原特异性)信号。通过表达信号调节蛋白-α(SIRPα;也称为SHPS1)的特定DC子集进行IL-6的转导可诱导IL-23R的表达,这对于致病性TH17细胞的扩增和稳定至关重要。除TH17细胞外,T卵泡辅助细胞(TFH)是依赖于IL-6生成的T细胞的第二个子集。但是,TFH细胞对IL-6的感应方式尚不清楚。此外,TFH细胞最有可能在两步过程中引发,该过程涉及DC介导的刺激和诱导CXC-趋化因子受体5(CXCR5),对重新定位到生发中心至关重要的趋化因子受体和转录因子B细胞淋巴瘤。6蛋白(BCL-6),它决定TFH细胞的转录特性。然后需要通过与B细胞的同源相互作用来稳定TFH细胞-这一过程除了诱导型T细胞共刺激物(ICOS)和ICOS配体(ICOSL;也称为ICOSLG)相互作用之外,还需要T细胞感应IL-6 。
4、IL-6与GC-TFH细胞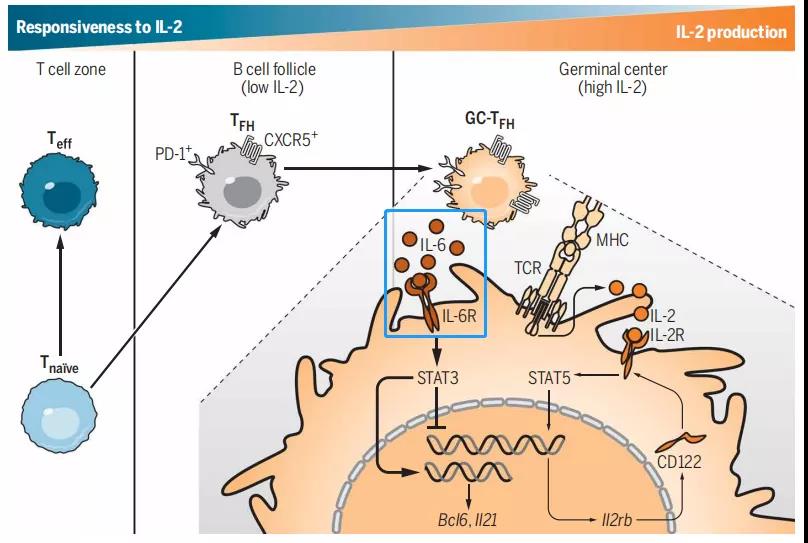
在流感病毒感染的小鼠模型中,TFH细胞中的IL-6信号激活STAT3,后者与IL-2诱导的STAT5竞争结合II2r基因座。通过抑制TFH细胞上的CD122(IL-2R)表达,IL-6降低了细胞对IL-2的反应性,并促进了GC-TFH细胞的生成。
五、IL-6在疾病中的作用
IL-6刺激多种疾病的炎症和自身免疫过程,例如糖尿病,动脉粥样硬化,抑郁症,阿尔茨海默氏病,系统性红斑狼疮,多发性骨髓瘤,前列腺癌,白塞病,和类风湿关节炎等。
1、IL-6信号通路及其在人类疾病发病机制中的作用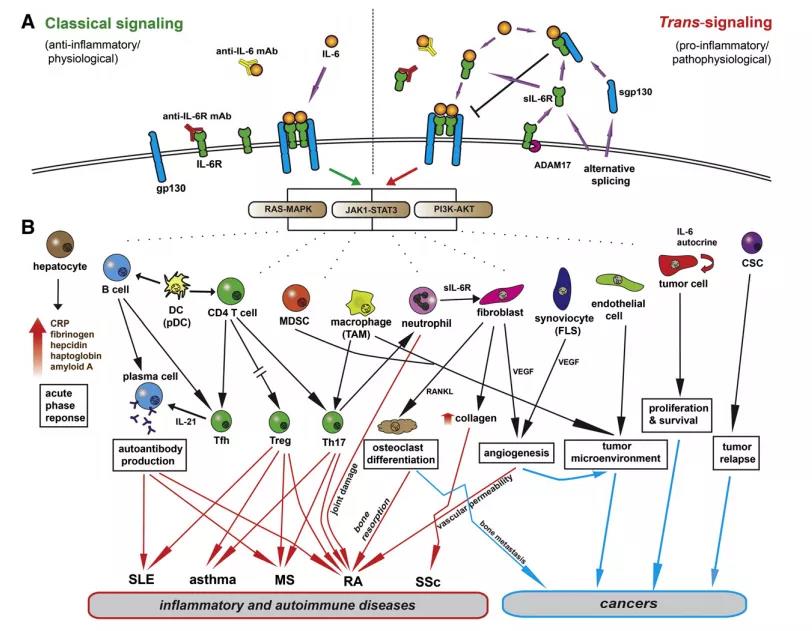
A.经典途径和反信号途径被认为分别有助于IL-6的抗炎和促炎活性。经典信号传导是由膜相关的IL-6R介导的,而可溶性信号形式的sIL-6R则是反式信号传导。sIL-6R是通过金属蛋白酶ADAM-17的切割或选择性剪接产生的,而sgp130仅通过选择性剪接产生。gp130在广泛的细胞类型中普遍表达,因此可通过反信号机制解释IL-6的多能活性。经典和跨信号通路均激活相同的细胞内通路,包括JAK1-STAT3,RAS-MAPK和PI3K-AKT。抗IL-6R mAb(红色)可阻断IL-6与膜结合的IL-6R和sIL-6R的结合。抗IL-6 mAb(黄色)与IL-6分子结合并抑制经典信号和反信号信号。sgp130充当IL-6反信号的诱饵抑制剂。
B. IL-6的生物学功能有助于炎症性自身免疫性疾病和癌症的发病机理。IL-6在炎症过程中激活肝细胞以释放急性期蛋白。IL-6由涉及自身免疫的多种活化细胞类型产生。活化的浆细胞样树突状细胞(pDC),常规树突状细胞(DC),B细胞和T细胞产生IL-6。IL-6是B细胞分化和血浆B细胞存活的重要细胞因子。这对于在淋巴组织的生发中心的T滤泡辅助细胞(Tfh)的活性至关重要,这对于通过IL-21产生自反应性B细胞至关重要。IL-6促进Th17分化,而相反地抑制CD4 + Foxp3 + T调节细胞(Treg)。因此,抑制IL-6信号传导在控制由T和B细胞介导的自身免疫和炎症方面具有深远的影响。另外,IL-6的炎性作用由其他细胞类型介导,包括巨噬细胞,嗜中性粒细胞,成纤维细胞,成纤维细胞样滑膜细胞(FLS)和内皮细胞。IL-6具有成纤维性,可诱导成纤维细胞产生胶原蛋白。它支持破骨细胞的分化并分别通过核因子κB配体(RANKL)和血管内皮生长因子(VEGF)的受体激活剂增强血管生成。IL-6通过旁分泌和自分泌机制促进肿瘤细胞的增殖和存活。在被IL-6激活后,各种类型的细胞(如肿瘤相关的巨噬细胞(TAM),髓样来源的抑制细胞(MDSC)和内皮细胞)参与促炎性和转移性肿瘤微环境的发展。另外,IL-6调节肿瘤微环境中Treg和Th17之间的平衡,并促进非癌症干细胞产生癌症干细胞(CSC)。
2、IL-6家族细胞因子与生理和疾病过程的联系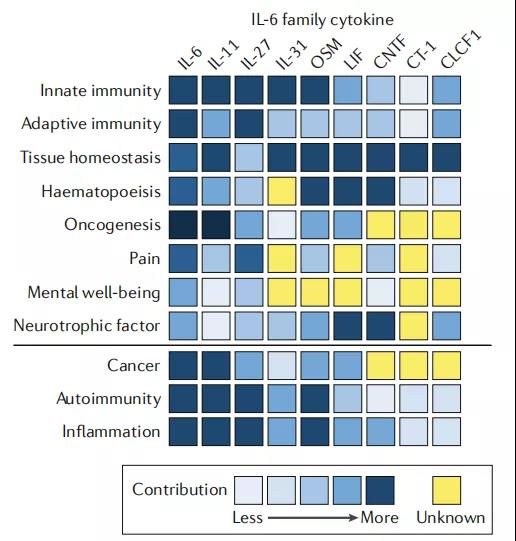
面板中描述的热图详细说明了单个IL-6细胞因子家族成员的情况 有助于特定的生理和免疫过程,并强调它们在某些疾病环境中的相对重要性。
3、IL-6与炎症,免疫力和疾病有关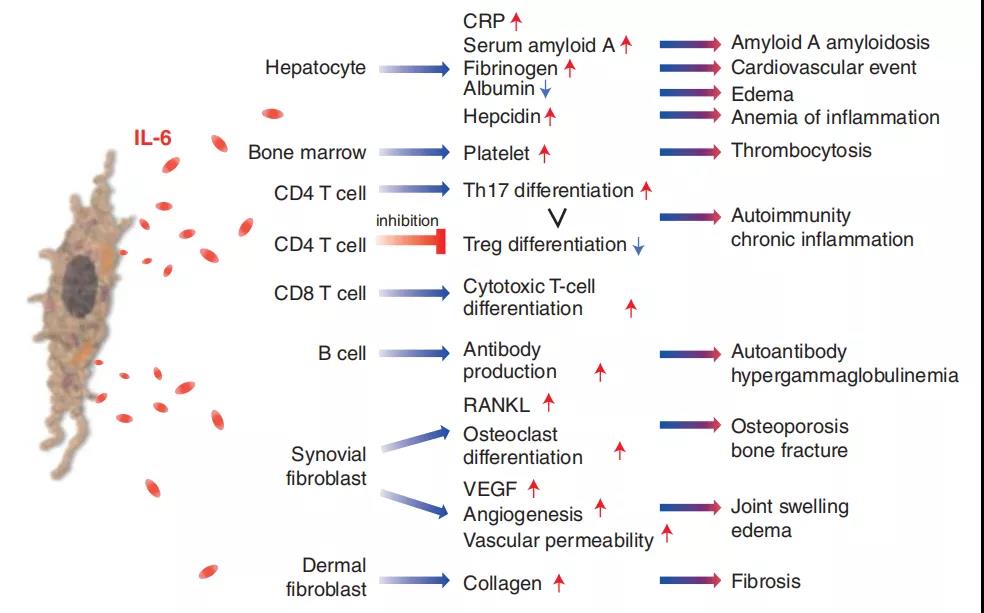
IL-6是具有多效活性的细胞因子。它诱导肝细胞中急性期蛋白的合成,例如CRP,血清淀粉样蛋白A,纤维蛋白原和铁调素,而它抑制白蛋白的产生。IL-6通过刺激抗体产生和效应T细胞的发展,在获得性免疫应答中也起重要作用。此外,IL-6可以促进几种非免疫细胞的分化或增殖。由于多效性活动,IL-6持续生产失调导致各种疾病的发作或发展。
4、IL-6在急性炎症中的作用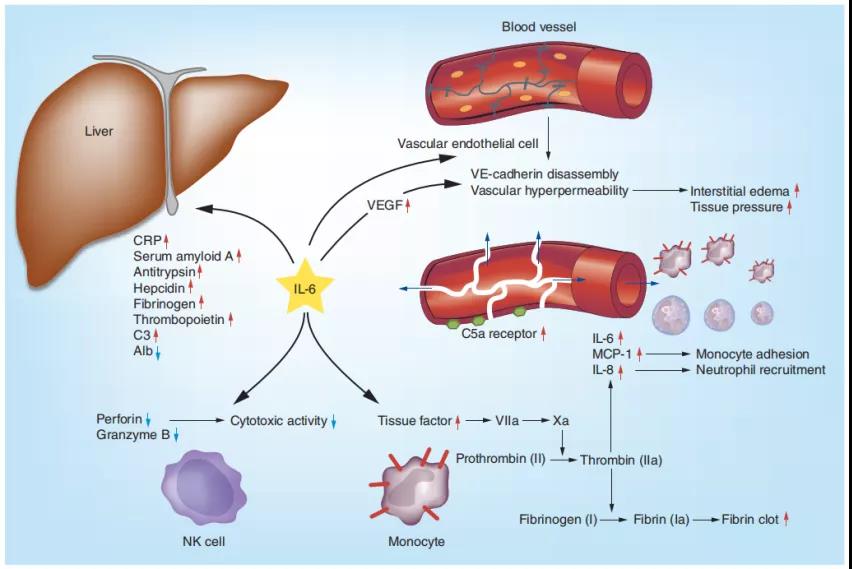
IL-6可从肝脏快速诱导多种蛋白,例如CRP,血清淀粉样蛋白A,抗胰蛋白酶,铁调素,纤维蛋白原,血小板生成素和补体C3。IL-6直接或通过诱导VEGF诱导内皮细胞上血管内皮钙粘蛋白的分解,从而导致血管通透性过高,从而导致组织损伤。IL-6上调内皮细胞上的C5a受体并增加其对C5a的反应性,从而进一步增强血管通透性。IL-6还诱导单核细胞表面上的组织因子表达并触发凝血级联反应,从而导致凝血酶活化和血纤蛋白凝块形成。凝血酶诱导内皮细胞上IL-6的表达,而IL-6增加MCP-1和IL-8的合成,从而触发单核细胞和中性粒细胞募集到血管内皮细胞。IL-6还可以通过减少穿孔素和颗粒酶B来抑制NK细胞的细胞毒活性。
5、IL-6对类风湿关节炎滑膜中骨代谢的影响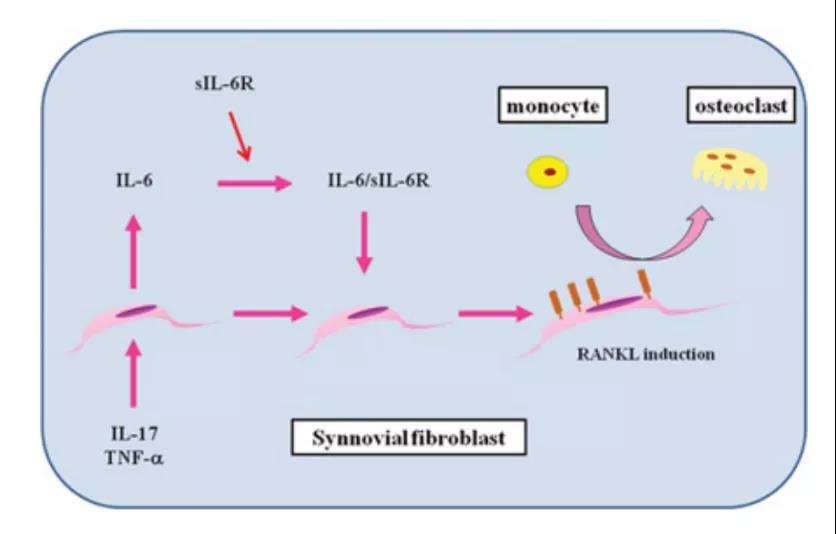
6、IL-6在RA发病机理中的各种作用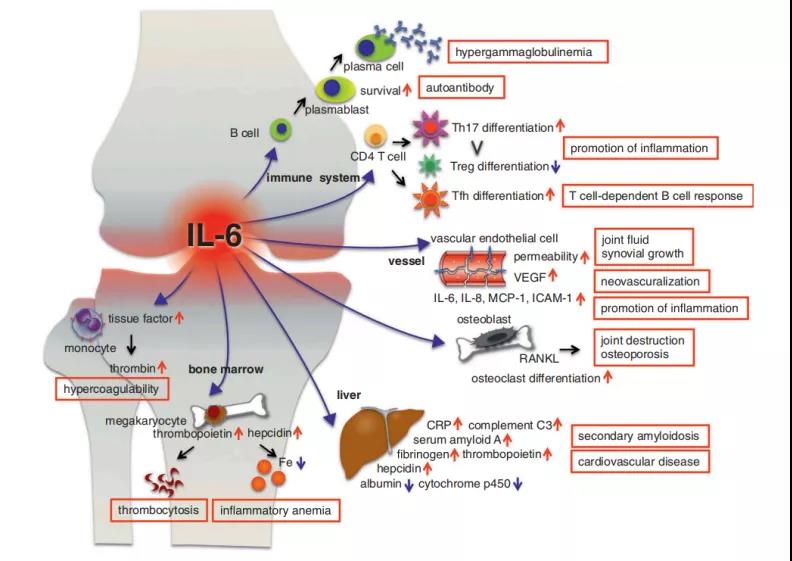
在RA患者中观察到的症状和并发症用方框标出。在免疫细胞中,IL-6促进B细胞分化为产生Ig的浆细胞,并支持浆母细胞的存活。浆细胞和成浆细胞产生抗瓜氨酸化的蛋白抗体。IL-6支持将原始CD4+ T细胞分化为Th17,并抑制其分化为Treg细胞,从而使Th17/Treg平衡朝着Th17优势转移并促进炎症。IL-6诱导Tfh细胞分化,这是依赖T细胞的B细胞反应所必需的。在受影响的关节中,IL-6通过诱导血管内皮生长因子(VEGF)直接或间接作用于血管内皮细胞,以促进血管渗漏,从而导致滑液积聚,关节肿胀和滑膜生长。IL-6刺激成骨细胞和滑膜细胞产生核因子(NF)-|B配体(RANKL)的受体激活剂,后者激活破骨细胞,导致骨质疏松和关节破坏。至于全身症状,IL-6迅速诱导急性期蛋白的产生。C反应蛋白(CRP),血清淀粉样蛋白A,补体C3,纤维蛋白原,铁调素和血小板生成素,但会降低白蛋白和细胞色素p450。持续升高的血清淀粉样蛋白A水平会导致继发性淀粉样变性。IL-6在肝脏中诱导铁调素,导致炎症性贫血。IL-6还促进巨核细胞的成熟并诱导血小板生成素,进而促进巨核细胞祖细胞的扩增和血小板的产生。纤维蛋白原水平升高和血小板增多与心血管疾病的风险有关。最后,IL-6诱导单核细胞上的组织因子,从而促进凝血。
7、IL-6是肝脏再生的主要驱动力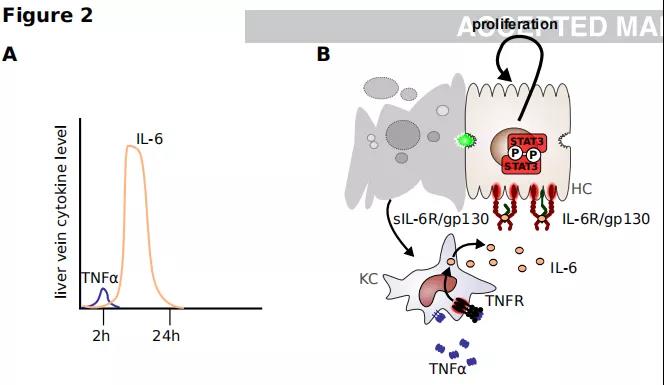
肝细胞损伤后的早期,库普弗细胞分泌TNFα,从而以自分泌方式诱导IL-6的大量表达。然后IL-6刺激肝细胞增殖。
8、IL-6对代谢的多重影响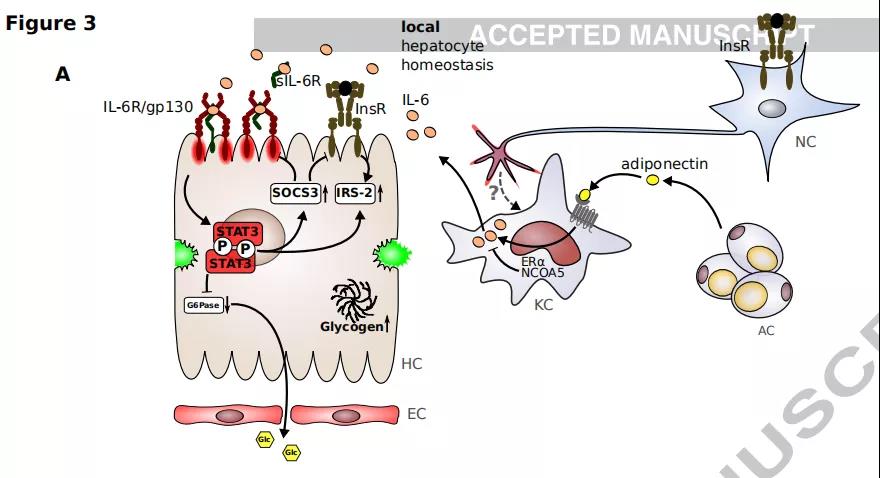

A.胰岛素受体(InsR)在表达刺豚鼠相关肽(AgRP)的神经元上的参与导致肝脏中IL-6的表达。Kupffer细胞对IL-6的表达受雌激素受体α(ERα)和核受体共激活因子5(NCOA5)负调节。IL-6在肝细胞上诱导胰岛素受体底物2(IRS-2)的表达,从而增强胰岛素信号传导。IL-6依赖性葡萄糖6磷酸酶(G6Pase)抑制可降低外周血葡萄糖水平并增加肝细胞中糖原的储存。IL-6对细胞因子信号传导抑制因子3(SOCS-3)的上调损害了gp130和InsR信号传导。
B.肝IL-6信号的丢失导致肝脂肪变性,局部炎症,血糖水平升高和由TNFα介导的外周胰岛素抵抗。
9、IL-6促进肝癌发生的多个步骤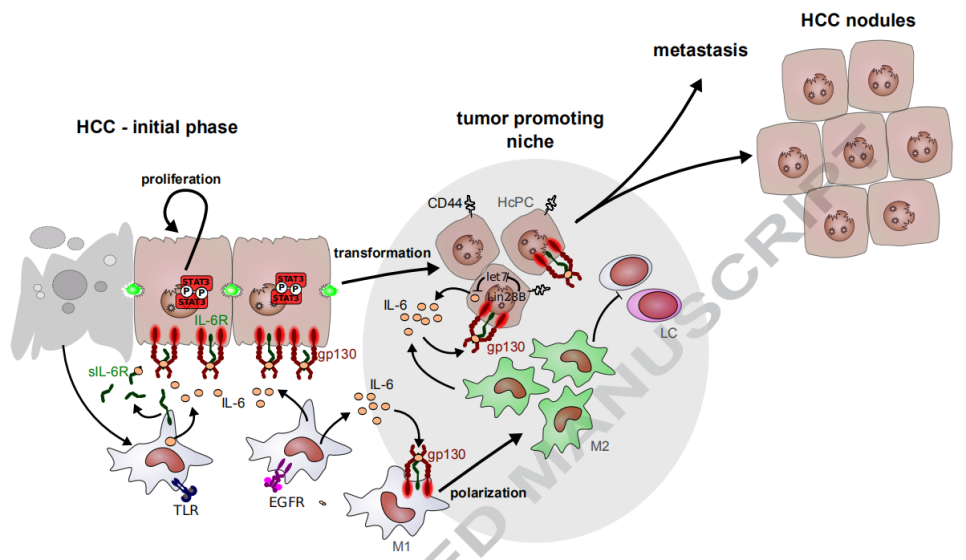
Kupffer细胞被受损的肝细胞以TLR和EGFR依赖性方式激活,并分泌IL-6和可溶性IL-6R,从而导致代偿性肝细胞增殖。在促进肿瘤的微环境中,肝细胞转化为HCC祖细胞(HcPC)通过LIN28B的上调获得自分泌IL-6回路。还分泌IL-6的M2型巨噬细胞促进肝细胞转化。IL-6促进M1到M2巨噬细胞极化。HcPC从促进肿瘤的小生境中流出,形成HCC结节,最终转移到远处的器官。
10、在疾病的背景下,IL-6可能具有局部炎症和全身作用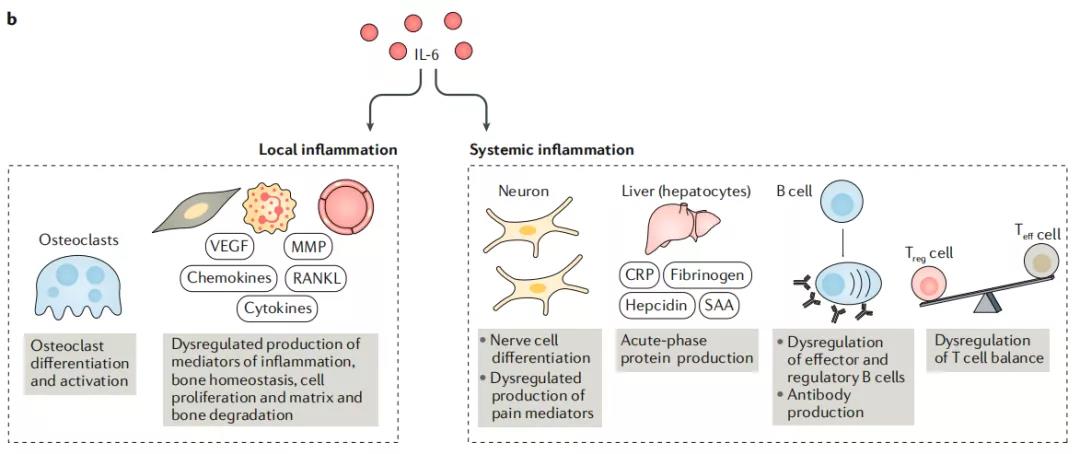
在临床前和临床数据的基础上,可以通过IL-6的作用来解释批准使用IL-6抑制剂的某些疾病表现。IL-6与疾病的发病机理有关,包括类风湿性关节炎,系统性幼年特发性关节炎(sJIA),Castleman病,巨细胞动脉炎,Takayasu动脉炎和细胞因子释放综合征。
11、iMCD发病机制与IL-6
三种假设的机制可能是iMCD细胞因子和趋化因子风暴的原因:首先,自身免疫/自身炎性假设涉及(1a)自身抗体通过抗原呈递细胞触发促炎性细胞因子释放,诱导目前尚不知道的分泌高细胞因子的细胞释放IL-6或其他病理细胞因子;(1b / c)抗原呈递细胞或其他迄今未知的分泌高细胞因子的分泌细胞释放IL-6或其他病理性细胞因子的信号失调,或(1d)激活的炎性细胞的调控缺陷。IL-6的正反馈使细胞因子和趋化因子风暴持续存在,其他病理性细胞因子和/或可能进一步的自身抗体刺激。
其次,副肿瘤综合症假说涉及淋巴结内或外的良性或恶性细胞的体细胞突变,引起体细胞因子的释放。初步数据表明,这些可能是淋巴结基质细胞。
第三,病原体假说涉及临床上无法检测到的HHV-8感染,新型病毒或另一种指示促炎性细胞因子的病原体。根据病原体发现研究产生的初步数据,单一病毒无法主动感染。不论病因如何,细胞因子和趋化因子风暴是导致iMCD随后临床和组织病理学特征的常见途径。
12、Il-6家族细胞因子在癌症中内在和外在的特性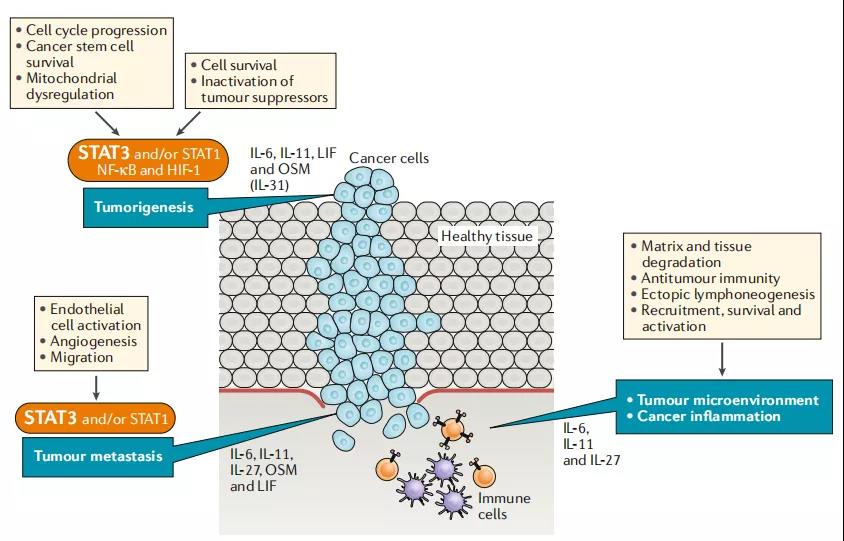
该图概述了在肿瘤发生和转移过程中,IL-6细胞因子家族成员调控的细胞过程,以及信号转导子和转录激活子3(STAT3)的主要激活。肿瘤微环境和癌症炎症。显示了与这些活动的控制有特定联系的细胞因子。注意,与肿瘤转移相关的过程也可有助于肿瘤微环境的整体结构,并支持与癌症炎症发展有关的活动。
六、IL-6细胞因子家族作为治疗靶标
IL-6细胞因子家族的成员是治疗感染,炎症,自身免疫和癌症的药物靶标。药物类别包括针对特定细胞因子或细胞因子受体的单克隆抗体和重组蛋白形式以及调节与膜糖蛋白130(gp130)或Janus激酶(JAK)相关的受体信号传导活性或细胞内机制的小分子干预–信号转导子和转录激活子(STAT)信号。显示了描绘IL-6细胞因子家族的代表性细胞因子细胞因子受体复合物的漫画。这些药物类别的示例显示在分区目标上。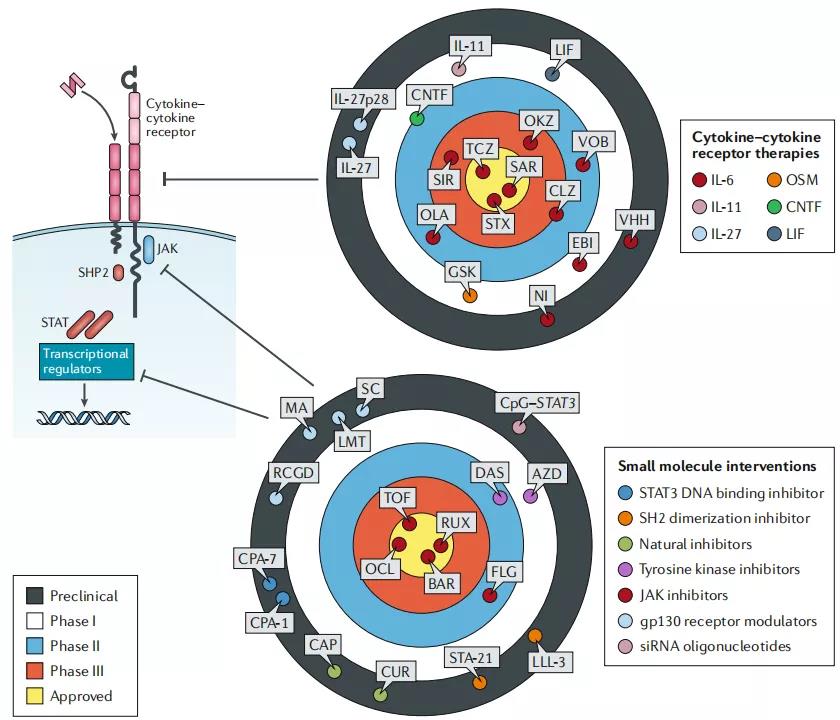
由于IL-6在免疫系统和炎症系统的功能障碍中具有多种作用,因此抗IL-6R治疗可以缓解各种症状,例如发烧,疲劳,疼痛,关节破坏,贫血等。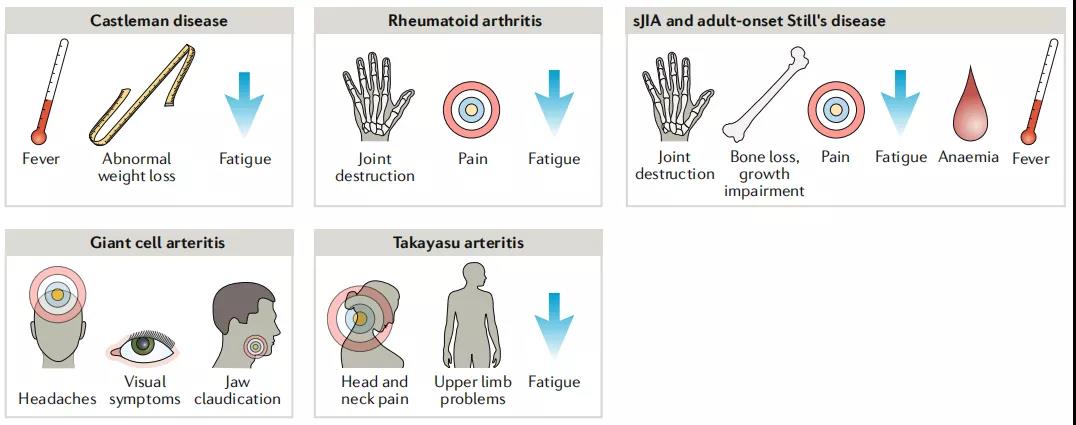
定量检测产品推荐: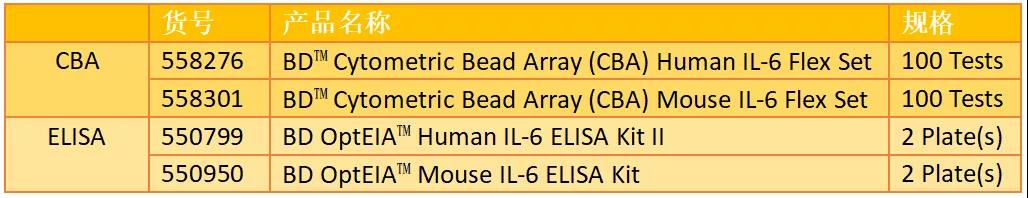
Reference:
[1].IL-6/IL-6 receptor system and its role in physiological and pathological conditions.
[2].IL-6: from its discovery to clinical applications.
[3].IL‑6 biology: implications for clinical targeting in rheumatic disease.
[4].IL-6 and the dysregulation of immune, bone, muscle, and metabolic homeostasis during spaceflight.
[5].The role and therapeutic targeting of IL-6 in rheumatoid arthritis.
[6].The role of IL‑6 in host defence against infections: immunobiology and clinical implications.
[7].IL-6 in Inflammation, Immunity, and Disease.
[8].Immunotherapeutic implications of IL-6 blockade for cytokine storm.
[9].IL-6 as a keystone cytokine in health and disease.
[10].Targeting interleukin-6 in inflammatory autoimmune diseases and cancers.
[11].Interleukin (IL-6) Immunotherapy.
[12].Interleukin-6: designing specific therapeutics for a complex cytokine.
[13].IL-6 pathway in the liver: from physiopathology to therapy.
[14].Translating IL-6 biology into effective treatments.
[15].Germinal center TFH cells: T(w)o be or not t(w)o be, IL-6 is the answer.
[16].Novel insights and therapeutic approaches in idiopathic multicentric Castleman disease.
[17].Recent insights into targeting the IL-6 cytokine family in inflammatory diseases and cancer.



 危险品化学品经营许可证(不带存储) 许可证编号:沪(杨)应急管危经许[2022]202944(QY)
危险品化学品经营许可证(不带存储) 许可证编号:沪(杨)应急管危经许[2022]202944(QY)  营业执照(三证合一)
营业执照(三证合一)