染色质重塑作为近年来国自然热点之一,虽然不像细胞死亡或者免疫调控那么热门,但是染色质的动态调控对于生物遗传具有重要的意义,所以染色质重塑经过这几年的发展,未来一定会是一个持续的研究热点。
(图片来源于网络)
△点击放大图片
今天我们通过两篇高分文章对于染色质重塑的研究进行分析,为大家后续的研究提供一些思路。
染色质重塑是和DNA甲基化和蛋白质修饰一样的可以对染色质进行影响变化的一种机体行为,但是因为一直不清楚染色质重塑的具体机制,所以研究进度较慢。直到近些年冷冻电镜的发展,让人们可以对染色质重塑的结构有了认识,从而可以对机制有了更深的研究。
例如BAF(BRG1-associated factors)和PBAF(polybromo-associated BRG1-associated factors)复合物是人类染色质调控因子,他们可以调控染色质结构变化从而影响基因表达。据目前研究发现,该复合物广泛参与动物细胞的发育分化过程,并且BAF/PBAF的突变会导致很多癌症及其他疾病。虽然科学界很早就发现鉴定了BAF/PBAF复合物,但是对于复合物的组合方式和对染色质重塑的影响机制一直不是很明确。正是近几年得益于冷冻电镜技术,酵母SWI/SNF家族复合物及人源BAF复合物的高分辨结构先后在2019和2020年得以报道。然而对于PBAF复合物的研究,结论尚少。
我们首先要看的这篇文章就是清华大学生命科学学院/结构生物学高精尖创新中心/清华-北大生命科学联合中心陈柱成教授研究团队在《自然》杂志在线发表的题为“Structure of human PBAF chromatin remodeling complex bound to a nucleosome”的研究论文。

△点击放大图片
陈柱成教授团队近年来一直致力于染色质重塑的研究,得到了很多研究成果,解释了染色质重塑的一部分机制。在之前的研究中,陈教授团队在酵母中发现了染色质重塑SNF2家族的马达蛋白是利用ATP水解产生的能量推动核小体的变化,从而实现对染色质的调控,了解到了基本的原理,此次更是突破技术难点通过体外重组的方法获得了高质量的PBAF复合物,并且报道了他们研究人源染色质重塑复合物PBAF在活性状态下结合核小体的结构,揭示了由12个亚基组成的PBAF复合物的组装方式,以及PBAF复合物识别核小体的机制,为后续人类疾病相关突变的致病机理提供了理论框架。
文章主要内容
01 PBAF复合体的整体结构
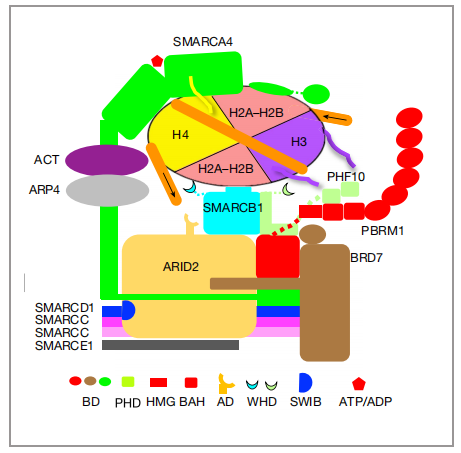
图1 与核小体结合的PBAF的结构示意图。箭头指示motor驱动的DNA易位的方向。ACT, β-actin; AD, arid domain; ARM, armadillo repeats; BD, bromodomain; CC, coiled coil; FH, finger helix.|图片取自文章
△点击放大图片
PBAF复合物的12个亚基可分为SMARCA4和11个辅助亚基(分别是:SMARCC1、SMARCC2、SMARCD1、SMARCE1、SMARCB1、ARID2、PHF10、PBRM1、BRD7、ARP4、ACTIN),他们分别有各自的功能,作者为了可以明确的把他们的功能区分开,把PBAF复合物的12个亚基,分为了3个模块(图2,a、b):1)发挥催化功能的马达(motor)模块2)调节肌动蛋白相关蛋白(ARP)模块3)染色质靶向模块(SRM)。
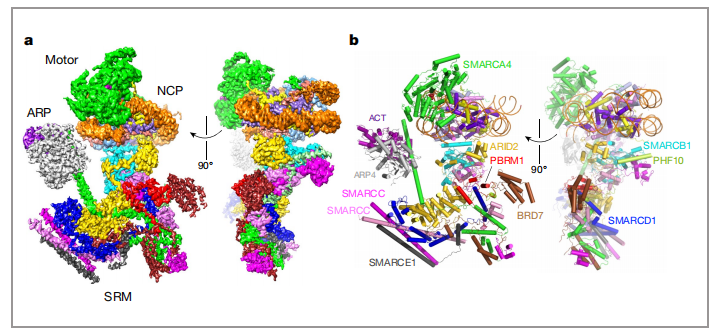
图2 a)人类PBAF与核小体结合的冷冻电镜结构图,包含核小体和结合核小体的3个部分(motor, ARP and SRM) b)PBAF的两个模型图 |图片取自文章
△点击放大图片
02 染色质识别SRM模块
PABF的大多数辅助亚基复杂的交织成负责识别核小体信号的SRM(图3,a),其中包含支架亚基SMARCC1和SMARCC2,转录因子结合亚基SMARCD1和SMARCE1,DNA结合亚基ARID2、组蛋白尾部结合亚基PBRM1、BRD7和PHF10,以及核小体结合亚基SMARCB1。并且他们形成明显的三叶状亚模块:核小体结合叶片(nucleosome-binding lobe,NBL)、组蛋白尾结合叶片(histone-tail-binding lobe,HBL)和DNA结合叶片(DNA-binding lobe,DBL)(图3,b)。其中结合组蛋白尾部的HBL是区别于BAF独有的一个亚模块,包含PBRM1、BRD7和PHF10(表1)。文章认为,这相当于一个超级组蛋白识别亚模块,有利于PBAF复合物在机体中更高效地感知染色质信号,发挥染色质重塑功能。文章分析了PBAF结合转录因子(TF)来调控基因表达的可能机理,同时也揭示了ARID2、PBRM1等亚基位点突变致病的可能机理。
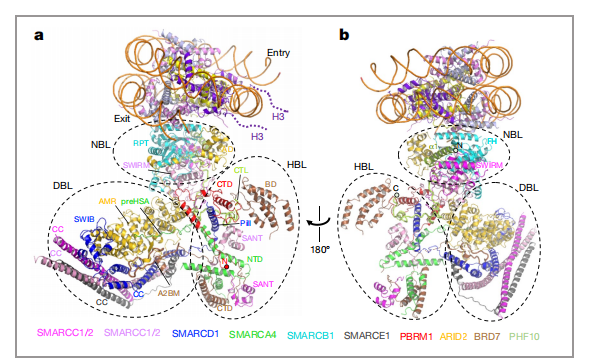
图3 a)和b)SRM的两个模型图,虚线椭圆代表参与核小体招募的三个叶状亚模块 |图片取自文章
△点击放大图片
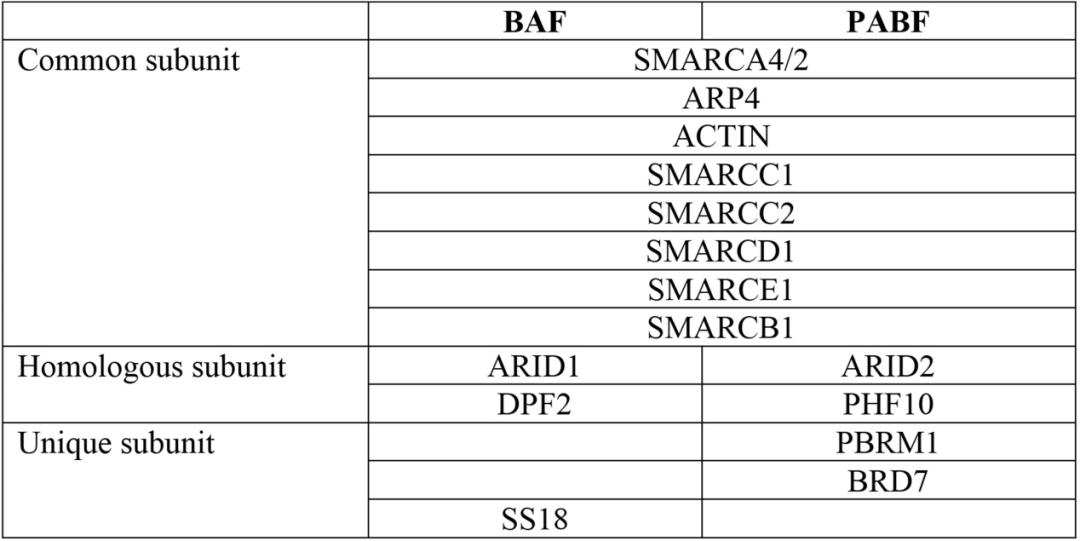
表1 BAF和PBAF的亚基差别 |表格取自文章
△点击放大图片
03 活性SMARCA4的马达结构
因为PBAF复合物的低分辨率结构和人BAF的高分辨率结构在之前已经被报道过了,所以本篇文章在此基础上,利用冷冻电镜技术,主要解析了PBAF结合核小体的高分辨率结构(3.4埃),并且在马达亚基SMARCA4的解析达到了2.8埃(图4),由此可以清楚的观察到催化ATP口袋和支撑螺旋(Brace-Helix)。支撑螺旋在染色质重塑中起到至关重要的作用,该结构介导了两个马达结构域(Lobe1与Lobe2)的相互作用,是耦合ATP水解和核小体滑动的关键(图5、6)。
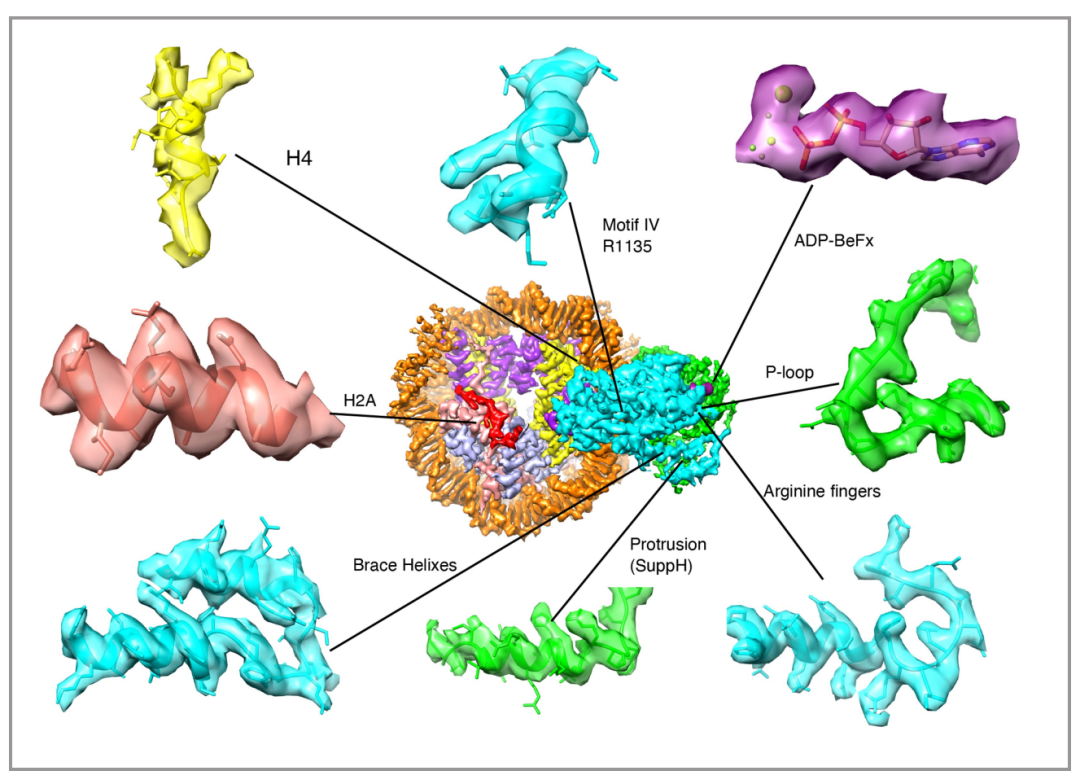
图4 分辨率为2.8埃的motor模型冷冻电镜图 |图片取自文章
△点击放大图片

图5 与核小体结合的motor结构域的局部图谱。Lobe1, lobe2和SnAc域分别显示为绿色、青色和红色 |图片取自文章
△点击放大图片
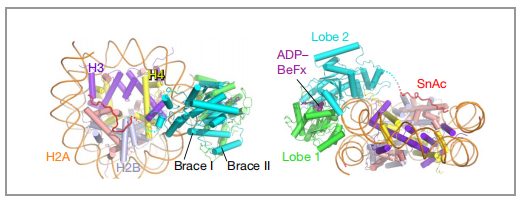
图6 与核小体结合的motor模型图 |图片取自文章
△点击放大图片
SMARCA4的运动结构域采用封闭构象,并与核小体紧密结合,这与之前报道的BAF的结构不同,说明了本研究解析的复合物中马达亚基SMARCA4处于活性状态。SMARCA4的活跃构象导致经常发生突变,然而我们可以将与疾病相关的突变映射到功能状态上。突变主要集中在两个位点上,一个在高度保守的ATP结合口袋周围,另一个围绕在新形成的支撑螺旋(Brace-Helix)周围。这些突变会导致染色质重塑活性明显降低,所以这些突变有可能导致PABF功能的丧失从而导致某些疾病。另外,高质量的电镜密度首次揭示了SMARCA4亚基的SnAc 结构域识别核小体的机制:SnAc通过3个精氨酸锚点识别核小体的酸性口袋,促进染色质重塑活性(图7)。
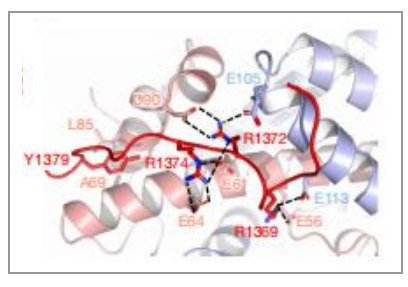
图7 SnAc结构域与核小体的酸性口袋之间的识别结合 |图片取自文章
△点击放大图片
总结
本篇文章不仅为PBAF复合物组装、核小体识别和染色质重塑机制提供了机制上的见解,也为PBAF复合体多个亚基经常发生突变导致的人类疾病机理提供了理论基础,更有利于后续理解染色质重塑研究,并且有可能推动相关疾病靶向药物的研发。
(因原文图片及细节描述过多,更多详情可查看原文章)
第二篇文章是来自美国密歇根大学转化病理学中心阿鲁·奇尼安(Arul M. Chinnaiyan)博士团队在 Nature 上发表了一篇研究论文,题为“Targeting SWI/SNFATPases in enhancer-addicted prostate cancer”,论文的共同第一作者是肖兰博博士。肖兰博博士在中南大学湘雅医学院曹亚教授实验室博士毕业后,在美国密歇根大学医学院的密歇根大学转化病理学中心(Michigan Center for Translational Pathology, MCTP)Arul M.Chinnaiyan 博士的团队完成了博士后研究,并作为 junior faculty 留校继续从事前列腺癌的基础研究。
△点击放大图片
该论文报道了一种针对 SWI/SNF 染色质重塑复合物 ATP 酶 SMARCA2 和 SMARCA4 的蛋白水解靶向嵌合体降解剂,称为 AU-15330。AU-15330能通过抑制增强子区域染色质的开放程度,高效抑制雄激素受体依赖型前列腺癌的生长,并与 AR 拮抗剂恩扎鲁胺协同作用,甚至在去势抵抗性前列腺癌(CRPC)模型中诱导疾病缓解。因此,阻碍 SWI/SNF 染色质重塑复合物介导的增强子可达性为增强子成瘾的癌症提供了一种有前途的治疗方法。
文章主要内容
01 AU-15330是靶向SWI/SNF染色质重塑复合物ATPase亚基的蛋白降解剂
AU-15330的结构图及与SMARC2和SMARCA4、PBRM1结合域如(图1a)。为了证明AU-15330降解剂的降解有效性,通过使用AU-15330处理多个细胞系后,导致的SMARCA2、SMARCA4和PBRM1的时间和剂量依赖性降解(图1b)。并且为了验证前列腺癌细胞对SWI/SNF ATP 酶降解剂AU-15330非常敏感,作者对来自14个不同谱系的正常和癌细胞系进行处理得出半抑制浓度(图1c),得出AU-15330能高效且选择性地抑制雄激素受体阳性前列腺癌细胞系的增殖。

图1:AU-15330是SWI/SNF ATP酶的特异性降解剂,在增强子结合转录因子驱动的癌症中表现出优先的细胞毒性 |图片取自文章
△点击放大图片
02 AU-15330抑制前列腺癌的作用机理
通过染色质开放性测序(ATAC-seq)分析了AU-15330对物理染色质可及性的影响。结果发现,在AU-15330处理一个小时后,VCaP细胞中超过30000个位点的染色质可及性迅速且接近完成丧失(图2a)。
通过染色质免疫沉淀测序(ChIP-seq)显示,在AU-15330处理后1小时内,VCaP细胞中AR、FOXA1和ERG的染色质结合减少(图2c)。作者还检测到H3K27Ac ChIP-seq信号中特征的“谷”模式的消失,表明侧翼核小体向AU-15330致密增强子的中心运动(图2d)。
通过RNA测序(RNA-seq)的全局转录组分析显示,在多个前列腺癌细胞中AR和foxa1调控基因显著下调,在ERG融合阳性VCaP细胞中ERG调控转录本显著下调,并且检测到AU-15330中MYC靶基因的表达显著缺失(图2e)。
综合,AU-15330是通过降解SWI/SNF染色质重塑复合物ATPase亚基快速降低前列腺细胞基因组部分区域(尤其是增强子)的染色质开放程度,阻断多个转录因子结合到其靶基因增强子区域的顺式作用元件上,从而显著抑制前列腺癌细胞中多条异常活化信号通路的活性。

图2:SWI/SNF ATP酶的降解破坏了核心增强子电路上的物理染色质可及性,从而使致癌转录程序失效 |图片取自文章
△点击放大图片
03 AU-15330抑制前列腺癌的作用机理2
在癌症中,AR、FOXA1和MYC等致癌基因的高表达已被证明是通过与多增强子簇的环状相互作用而连接的,通常被称为超级增强子。文章在AR、MYC和TMPRSS2-ERG基因的顺式作用中发现了几个这样的调控簇(图3b),经过AU-15330处理后导致这些位点立即压缩,并导致超级增强子的H3K27Ac、AR和FOXA1 ChIP-seq信号丢失(图3c);同时进行了H3K4me3(活性启动子标记)和H3K27Ac Hi-C结合ChIP-seq(HiChIP-seq)的对超级增强子和启动子进行检测,结果SWI/SNF失活显著破坏了顺式增强子与AR基因启动子的三维环状相互作用(图3d),也就说SWI/SNF atpa酶失活特异性地导致前列腺癌细胞中AR、FOXA1、ERG和myc激活的核心增强子电路的全基因组崩溃,从而恢复正常生理水平。
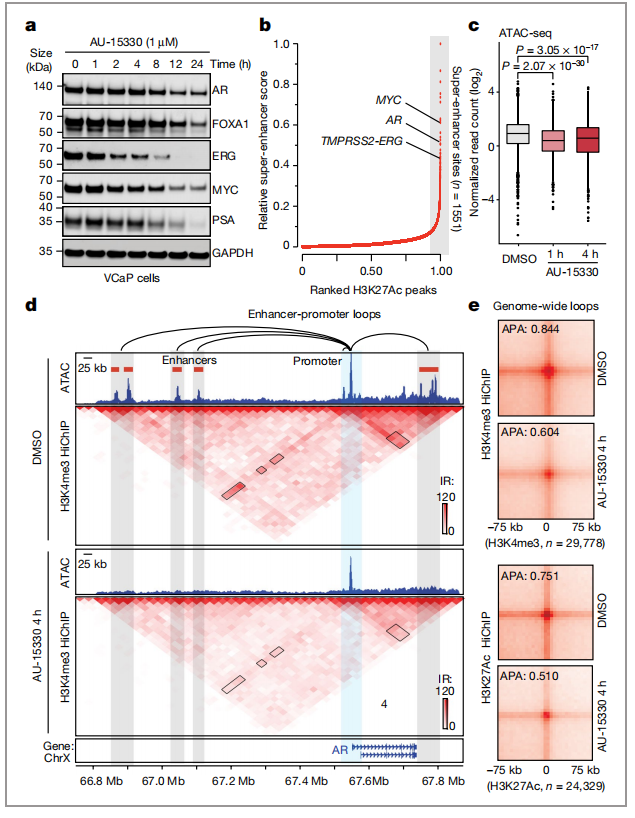
图3:SWI/SNF ATP酶降解破坏增强子-启动子环,以调节驱动致癌基因的超生理表达 |图片取自文章
△点击放大图片
04 AU-15330的安全性和有效性评估
通过对晚期前列腺癌动物模型中的AU-15330进行了药理学表征测试,评估AU-15330的安全性和有效期。
文章采用VCaP去势耐药前列腺癌(CRPC)模型(VCaP-CRPC)来评估AU-15330的疗效。用恩扎鲁胺(一种AR拮抗剂)治疗携带VCaP-CRPC异种移植物的阉割雄性小鼠显示出中等的抗肿瘤疗效;然而,用AU-15330治疗可有效抑制肿瘤生长,在超过20%的动物中引发疾病消退(图4a,b)。联合方案(AU-15330+恩扎鲁胺)治疗诱导了最有效的抗肿瘤作用,在所有实验动物中均有效(图4a、b)。
无论是单独使用还是使用恩扎鲁胺,在AU-15330治疗5天后,肿瘤显示SWI/SNF靶点和AR、ERG靶点、MYC和Ki67显著下调(图4c)。AU-15330也能强烈抑制C4-2B细胞系来源的CRPC异种移植物在完整小鼠体内的生长,并与恩扎鲁胺协同作用(图4d)。
组合方案还显著抑制了MDA-PCa-146-12的肿瘤生长,这是一种患者来源的异种移植(PDX)模型,固有地对恩扎鲁胺具有耐药性。通过将肿瘤植入去势小鼠体内,进一步建立了MDA-PCa-146-12 PDX的CRPC变体。即使在这种高度侵袭性的模型中,组合方案也诱导了显著的肿瘤生长抑制,导致超过30%的动物疾病退化(图4e)。在这些研究的所有两组中,我们都没有检测到动物体重的变化。在AU-15330治疗的小鼠中,胃肠道中也没有杯状细胞消耗的迹象,生殖细胞成熟没有缺陷,也没有睾丸萎缩,安全性得到了验证。
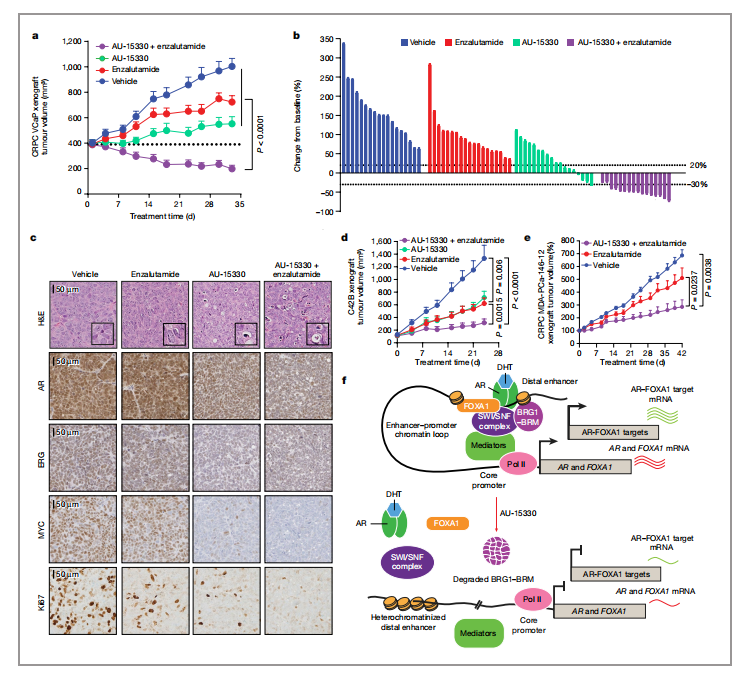
图4:AU-15330在CRPC的临床前模型中抑制肿瘤生长,并与恩扎鲁胺协同作用 |图片取自文章
△点击放大图片
综上所述,AU-15330可以特异性降解SWI/SNF染色质重塑复合物ATPase可能是一种新的前列腺癌表观遗传治疗策略。
部分相关优宁维可提供产品推荐:
| 货号 | 品名 | 应用 |
| SH30022 | DMEM with High Glucose, with 4.0 mM L-Glutamine, without Sodium Pyruvate | 细胞培养 |
| P3 Primary Cell 4D X Kit L | V4XP-3024 | 细胞转染 |
| 29401317 | StrepTrap XT 1X1ML | 蛋白纯化 |
| 29275878 | Capto HiRes Q 5/50 | 蛋白纯化 |
| 231222 | QIAGEN PCR Cloning plus Kit (10) | PCR |
| 11956 | SMARCC1/BAF155 (D7F8S) Rabbit mAb | WB |
| 52251 | Brg1 (E9O6E) Mouse mAb | WB |
| 5605 | c-Myc (D84C12) Rabbit mAb | WB |
| 8173 | Acetyl-Histone H3 (Lys27) (D5E4) XP® Rabbit mAb | WB |
| 9751 | Tri-Methyl-Histone H3 (Lys4) (C42D8) Rabbit mAb | WB |
| 9541 | Cleaved PARP (Asp214) Antibody (Human Specific) | WB |
| 97249 | ERG (A7L1G) Rabbit mAb | WB |
| 3418 | CTCF (D31H2) XP® Rabbit mAb | WB |
| 58214 | BAF Complex Antibody Sampler Kit II | WB |
| 12854 | BAF Complex Antibody Sampler Kit | WB |
| 78779 | PBAF Complex Antibody Sampler Kit | WB |
| 3508 | Brg1 (A52) Antibody | WB |
| 9002S | SimpleChIP® Enzymatic Chromatin IP Kit (Agarose Beads) | ChIP |
| 9003S | SimpleChIP® Enzymatic Chromatin IP Kit (Magnetic Beads) | ChIP |
| 9004S | SimpleChIP® Plus Enzymatic Chromatin IP Kit (Agarose Beads) | ChIP |
| 9005S | SimpleChIP® Plus Enzymatic Chromatin IP Kit (Magnetic Beads) | ChIP |
| 56383S | SimpleChIP® Plus Sonication Chromatin IP Kit | ChIP |
| 86652S | CUT&RUN Assay Kit | CUT&RUN |