酶学研究一直是热门领域研究的重点,近几年相关文章也是不断飙升。酶是生命活动中的“催化剂”,主宰着生命活动的进程。酶学研究对于人们认识生命活动的本质和规律是十分重要的,酶学研究中重大发现的理论上的突破也多次获得诺贝尔奖桂冠。
今天我们看到的这篇研究是来自美国费城威斯达研究所的Kazuko Nishikura教授2021年3月发表在Nat Commun上面的文章。Kazuko Nishikura教授致力于RNA编辑的研究,也是最早在哺乳细胞中发现ADAR1的研究者。这篇文章展现了Kazuko Nishikura对ADAR1 RNA编辑酶在癌症细胞中调节R-loop的形成和基因组端粒稳定性中的新发现。
文章条理清晰,数据详细扎实,非常适合于研究学习。文章中也用到了多种实验方法,如果大家对某些实验方法感兴趣,可以去看文章的补充材料或者找小优咨询哦!

△点击放大图片
ADAR1 RNA编辑酶在癌症细胞中调节R-loop的形成和基因组端粒稳定性
ADAR是一个作用于腺苷到肌苷(A-to-I RNA editing)的RNA编辑过程的重要的酶。ADAR1具有两个亚型,p150和p110。P150主要存在于细胞质,而P110主要存在于细胞核。前期的研究已表明P150介导的IFN信号通路抑制降低了肿瘤对免疫检查点阻滞的响应,这篇文章作者对p110进行进一步研究。

ADAR1作用以及上下游通路示意图
△点击放大图片
01 ADAR1的缺失导致DNA的损伤和端粒异常
首先故事开始于作者发现ADAR1基因敲除的HeLa细胞的延时摄影显示有异常形状的细胞核出现,在有丝分裂过程中发生阻滞(Fig A)。进一步检测发现,这些细胞中桥核,微核和多核显著增加。根据这些观察到的现象,作者猜测ADAR1基因和DNA损伤之间可能存在显著影响(Fig B)。

△点击放大图片
a,Hela细胞用siControl或者siADAR1进行转染,然后用CellLight Tubulin-GFP处理,细胞核位置通过用SIR-DNA试剂对DNA染色。
b,至少200个siControl或者siADAR1转染的Hela细胞用来检测细胞核异常。
作者进一步使用端粒FISH和γH2AX免疫染色发现ADAR1敲除的HeLa细胞中,端粒异常引起的变异损伤增多(黄色箭头指示)(Fig C)。
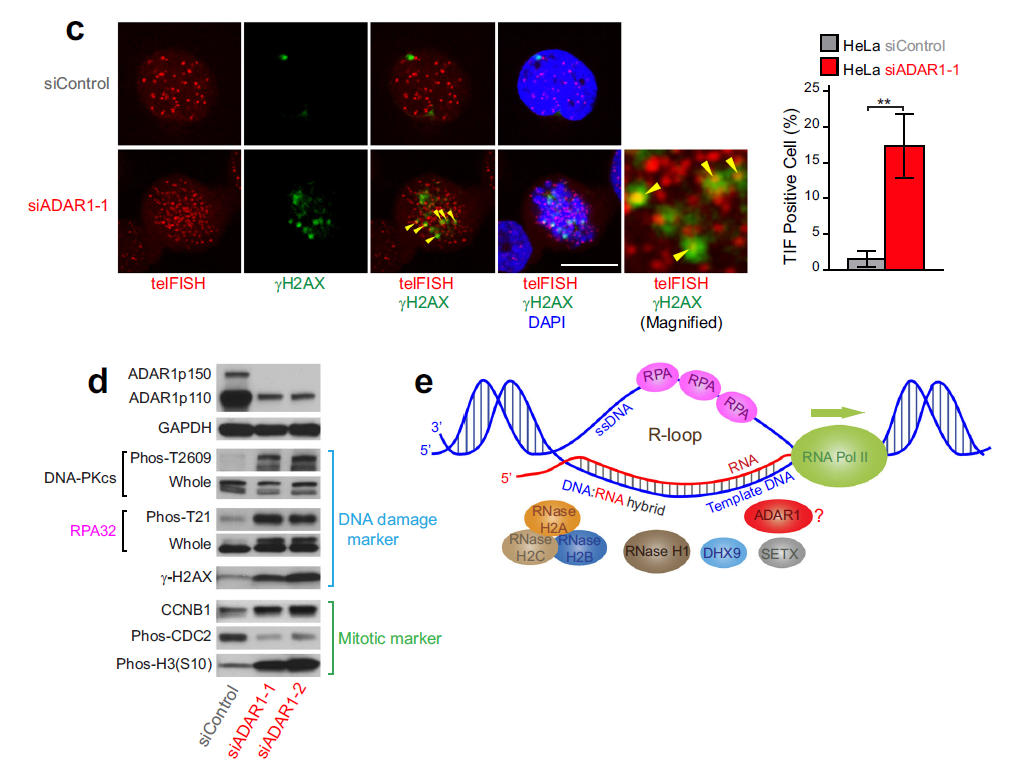
△点击放大图片
c,ADAR1敲除的细胞中端粒DNA损伤。端粒FISH和γH2AX免疫染色显示端粒异常导致的焦点显著增加(TIF,黄色箭头标记),表明在ADAR1敲除的HeLa细胞中,端粒重复DNA损伤和染色体异常有关。
WB实验显示DNA损伤标志物例如DNA-PKcs,H2AX和磷酸化的RPA32在ADAR1敲除的细胞中显著上调(Fig D)。作者猜测ADAR1的缺失可能会导致R-Loops的聚集(Fig E),从而会引起DNA的损伤,端粒异常和有丝分裂阻滞,从而开始第二部分的研究。

△点击放大图片
d,用siControl和siADAR1 RNA转染的Hela细胞提取物通过WB检测。
e,R-loop结构包含一个RNA聚合酶II在RNA链中形成的RNA:DNA杂交。
相关产品
| 货号 | 品名 | 规格 | 产品分类 |
| 10600023 | PVDF 0.45um | 30cm*4m | PVDF膜 |
| 10600002 | NC 0.45um | 30cm*4m | NC膜 |
| RPN2232 | ECL Prime WB detection reagent | 50ml+50ml | ECL发光液 |
| abs9301/abs9302 | 预制胶10%、12%,10、15 wells | 5片/盒 | 预制胶 |
| 4602 | DNA-PKcs Antibody | 10Oul | 一抗 |
| AF6654-SP | Anti-RPA32(Phos-T21) | 25ug | 一抗 |
| 52448 | RPA32/RPA2Antibody | 100ul | 一抗 |
| 4135 | Anti-Cyclin B1 (CCNB1) | 10Oul | 一抗 |
| 4539 | Anti-Phos-CDC2 | 100ul | 一抗 |
| 3377 | Anti-Phos-H3(S10) | 10Oul | 一抗 |
| abs20003 | Goat anti-Mouse IgG-FITC | 100ul | 荧光二抗 |
| Abs20004 | Goat anti-Rabbit IgG-FITC | 100ul | 荧光二抗 |
| 114-15 | jetPRIME | 1.5ml | siRNA转染试剂 |
| L-008630-00-0005 | ON-TARGETplus Human ADAR siRNA | 5nmol | siRNA |
| RHS4531一EG103 | GIPZ Lentiviral Human ADAR shRNA | 5nmol | shRNA |
| CM-008630-01-0002 | Edit-R CRISPR (knockout) Human ADAR crRNA | 2nmol | crRNA |
02 ADAR1缺失显著增加RNA:DNA杂交的形成
基于以上观察,作者开始了第二部分研究。作者进行了斑点实验发现ADAR1的缺失会显著增加RNA:DNA杂交的形成(Fig A and B)。使用Escherichia coli-RNase H(用于消化RNA链中RNA:DNA杂交链)会消除斑点信号(Fig B)。而ADAR2缺失的对照实验则显示ADAR2对于RNA:DNA杂交几乎没有影响(Fig C),显示ADAR1对R-Loop的特异性调节。

△点击放大图片
a,S9.6抗体识别特异性的RNA:DNA而不是DNA:DNA或者RNA:RNA核苷酸对。
b,S9.6抗体信号被E.coli-RHase H处理后破坏,确认RNA:DNA杂交的特异检测。
除了ADAR2的对照以外,作者还使用了酶活位点缺失的ADAR1突变体进行进一步对照实验,这也是酶活研究的常用方法。使用慢病毒表达体系进行转染,ADAR1基因的过表达显著抑制了RNA:DNA杂交的形成,而突变体ADAR1p110-E912A不能抑制杂交的形成(Fig E)。

△点击放大图片
e,内源ADAR1敲除之后通过转染ADAR1野生型而不是ADAR1酶活位点突变型造成RNA:DNA杂交的形成增加。
相关产品
| 货号 | 品名 | 规格 | 产品分类 |
| 13343 | Blood & Cell Culture DNA Midi Kit | 25Test | DNA提取试剂盒 |
| 28904276 | illustra tissue Mini | 250Test | DNA提取试剂盒 |
| abs9265 | proteinase K | 1ml | 蛋白酶K |
| 10262 | QIAGEN-Genomic-Tip 500/c (10) | 10Test | |
| RPN303B | HYBOND-N+ | 30cmk3m | 带正电尼龙膜 |
| abs20001 | Goat anti-mouse IgG-HRP | 100ul | 酶标二抗 |
| abs20002 | goat anti-rabbit IgG-HRP | 100ul | 酶标二抗 |
| 115-0015 | PEIpro | 1.5ml | 慢病毒转染试剂 |
03 ADAR1敲除引起RNA:DNA杂交在端粒重复区域聚集增加
作者接下来进一步研究ADAR1p110是如何控制R-loop的形成。作者对DNA:RNA杂交免疫共沉淀产物(DRIP)进行定量PCR,结果表明,ADAR1在已知的转录起始位点例如NEAT1,JUN,PMS2和CLSPN都没有显著作用。看来事情没有预想这么简单,作者进一步查阅文献,发现着丝粒和端粒重复区域也被报导对R-Loop的形成具有显著影响,标志基因是Alu和LINE。于是作者将DRIP产物又进行了斑点杂交试验,结果表明ADAR1敲除会引起RNA:DNA杂交在端粒重复区域聚集增加(Fig B)。

△点击放大图片
b.DRIP产物通过基因组的DNA斑点杂交实验进行检测。ADAR1敲除导致端粒重复序列上RNA:DNA的杂交形成增加,会被RNase H处理破坏。
04 ADAR1促进RNase H2酶对端粒R-loop识别
已知ADAR1是一个作用于腺苷到肌苷(A-to-I RNA editing)的RNA编辑过程的重要的酶,而在前面的研究又发现ADAR1的对于RNA:DNA在端粒重复区杂交聚集有影响,这个影响和ADAR1本身的功能又是什么关系呢?作者对于这个问题又进行了进一步的更加细致的机制研究。
作者猜测RNA:DNA杂交包含A-C错配碱基对可能有两形成途径,变体TCAGGG和TTGGG(Fig A and B)。
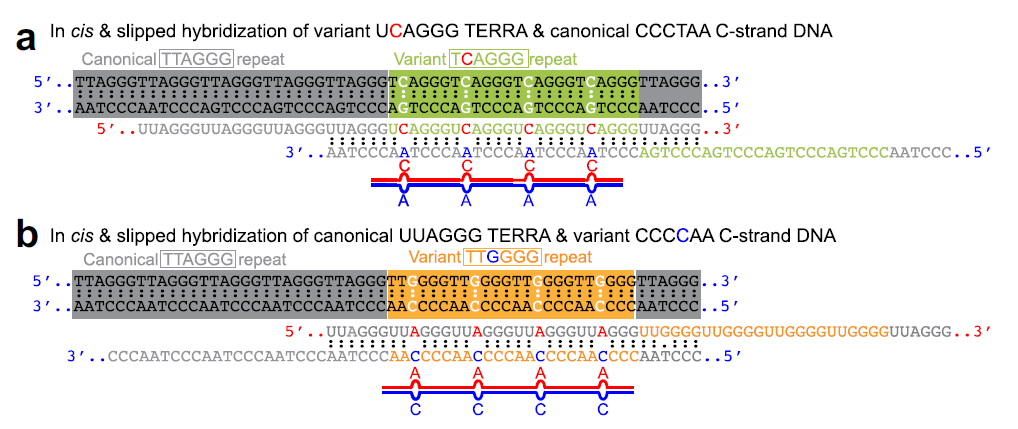
△点击放大图片
a,b RNAs transcribed from the region containing four TCAGGG (green) / TTGGGG (orange) variant repeats surrounded by TTAGGG (gray) canonical repeats form an RNA:DNA hybrid containing four A-C mismatches by in cis slipped hybridization to the C-strand DNA containing canonical TTAGGG (CCCTAA) /TTGGGG (CCCCAA) repeats.
作者发现,ADAR1的缺失会引起这两种RNA:DNA的聚集(Fig C and D)。

△点击放大图片
c. DRIP产物通过斑点实验检测UCAGGG变体和UUAGGG重复G-strand RNAs。
d. DRIP产物通过斑点实验检测CCCTAA变体和CCCCAA重复C-strand RNAs。
接下来,作者纯化ADAR1蛋白进行体外实验。作者使用了昆虫表达体系,用构建好杆状病毒载体感染Sf9昆虫细胞,从而获得表达的重组蛋白。然后作者使用了ni 填料进行纯化。体外实验也表明,ADAR1可以编辑端粒RNA:DNA杂交中的A-C错配。
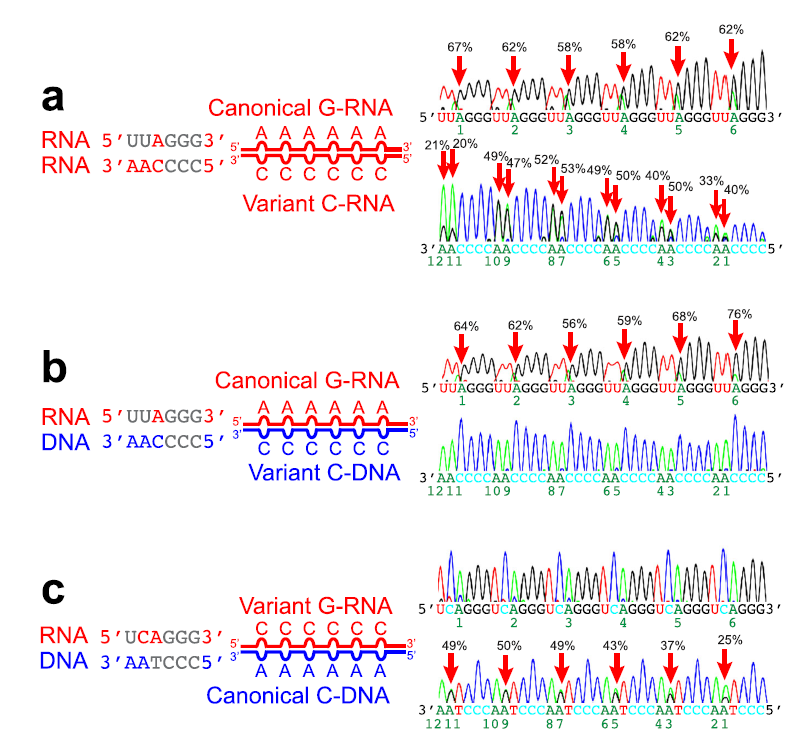
△点击放大图片
a,b,c. 使用重组ADAR1蛋白构建的包含A-C或者C-A错配的端粒重复序列(a:dsRNA,b,c:RNA:DNA杂交)体外编辑实验。PCR产物使用Sanger测序。
而对于端粒重复的RNA:DNA杂交中A-C错配的编辑又促进了他们被RNase H2酶识别。实验表明,RNase H2不能降解端粒重复RNA:DNA杂交中含有六个A-C错配的RNA链,开始降解包含四个A-C错配的RNA链,而用I:C代替所有的A-C错配之后可以引起RNase H2酶的高效降解。

△点击放大图片
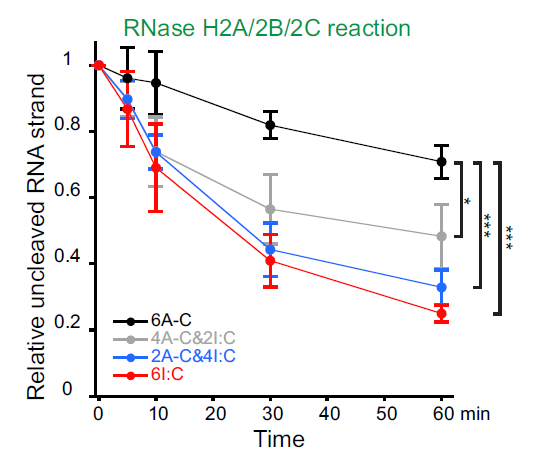
△点击放大图片
b,RNase H2不能降解包含6个A-C错配的端粒重复的RNA:DNA杂交的RNA链,但是开始降解包含4个A-C错配的端粒重复的RNA:DNA杂交的RNA链。用I:C匹配的碱基对替换所有的A-C错配导致RNAse H2对RNA链的高效降解。
相关产品
| 货号 | 品名 | 规格 | 产品分类 |
| 17531806 | NI SEPHAROSE 6FF ,5ML | 5ml | his标签填料 |
| 17531801 | NI SEPHAROS 6FF,25ML | 25ml | his标签填料 |
| 17075601 | GLUTATHIONE SEPHAROSE 4B ,10ML | 10ml | GST标签填料 |
| 17075605 | GLUTATHIONE SEPHAROSE4B,100ML | 100ml | GST标签填料 |
综上,作者发现了在癌症细胞中ADAR1可以调节端粒上R-loop的形成和基因组稳定性。ADAR1编辑RNA:DNA杂交中的A-C错配变为I:C配对,从而促进RNase H2酶对端粒R-loop的识别。由此表明ADAR1可以作为抗肿瘤免疫研究的有价值的治疗靶点。
就像这篇文章一样,在酶研究中我们常常会需要用到纯度较高的酶进行体外实验,构建并纯化出一个我们需要的酶是实验的关键。优宁维为您提供从质粒构建,蛋白表达到蛋白纯化全套解决方案。