自2018年肿瘤免疫获得了诺贝尔生理学或医学奖以来,肿瘤免疫一直是研究的大热门,在国自然基金的申请和中标也增量客观。肿瘤免疫的研究可以从肿瘤微环境,肿瘤免疫逃逸机制,免疫检查点,肿瘤免疫相关通路进行研究。
今天小编就为大家带来一篇来自耶鲁大学Lieping Chen教授在2019年发表在Cell上面的文章“Fibrinogen-like protein 1 is a major immune inhibitory ligand of LAG3”,文章逻辑清晰,论证缜密,希望能为大家带来新的思路。文章干货较多,建议收藏哦!
LAG3(Lymphocyte-activation gene 3)是一个主要在活化的T细胞上表达的跨膜蛋白。LAG3受到IL-2和IL-12的激活,作为一个受体来传递抑制性信号。LAG3可以抑制性的调节CD8和CD4 T细胞的增殖,激活,效应功能和稳态。本篇文章作者通过研究发现,纤维蛋白原样蛋白1(fibrinogen-like protein 1,FGL1),一个肝脏分泌蛋白,是一个不依赖于MHC-II的LAG3的主要的功能性配体。FGL1抑制抗原特异性T细胞激活,FGL1沉默会促进T细胞免疫。此外,血浆中FGL1高表达和抗PD-1/B7-H1治疗不良预后有关。作者的研究表明了一个免疫逃逸新机制和癌症治疗的新应用。
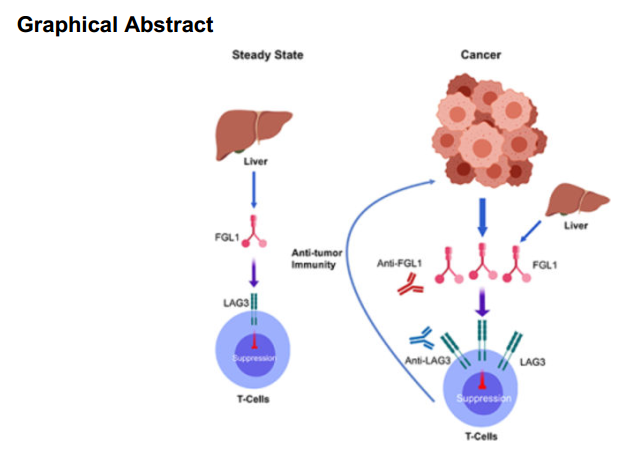
Graphical Abstract
△点击放大图片
PART1 FGL1是一个LAG3高亲和性的配体
作者使用了GSRA(Genome-Scale Receptor Array)技术来寻找LAG3的结合蛋白,通过Fc标记的LAG3胞外结构域进行筛选。结果表明FGL1是LAG3的主要结合蛋白(Fig.1a and Fig.1b)。作者用流式和BIAcore进一步验证,也表明FGL1和LAG3有相互作用。FGL1/LAG3相互作用的亲和力为1.5nM(Fig.1c and Fig.1d)。

Fig.1 a-d FGL1是一个LAG3的配体蛋白
△点击放大图片
FGL1包含了两个结构域,coil-coil domain(CCD)和fibrinogen-like domain(FD)。通过结构域突变研究,作者发现FD,而不是CCD结构域和LAG3有相互作用(Fig. S1e)。LAG3蛋白包含4个Ig-like胞外结构域,D1-D4。D3-D4结构域的去除不影响FGL1的结合,而D1或者D2结构域的分别去除会部分降低结合,表明D1和D2结构域对于FGL1/LAG3的结合都发挥重要作用(Fig. 1e)。
之前的研究表明LAG3 D1结构域的C端单点突变(Y73F)会干扰MHC-II结合。然而,该位点突变不会影响FGL1的结合,表明FLG1/LAG3的相互作用和MHC-II/LAG3的结合无关(Fig. 1e)。LAG3过表达的293T细胞预孵育C9B7W(LAG3的中和抗体,可以结合LAG3的D2结构域而不影响FGL1/MHC-II相互作用),引起FGL1/LAG3的相互作用完全中止(Fig. 1f)。LAG3过表达的细胞加入MHC-II融合蛋白也没有降低FGL1/MHC-II相互作用(Fig. S1f)。

Fig.1 e-f FGL1是一个LAG3的配体蛋白
△点击放大图片

Fig.S1 e-f FGL1/LAG3相互作用研究
△点击放大图片
综上表明,FGL1和LAG3的相互作用不依赖于MHC-II,而且这个相互作用涉及到FGL1的fibrinogen-like结构域和LAG3的D1-D2结构域。
BIAcore相关产品
| 货号 | 名称 | 规格 |
| BR100530 | Series S Sensor Chip CM5 | 3片装 |
| 29149603 | Series S Sensor Chip CM5 | 10片装 |
| BR100050 | Amine Coupling Kit | / |
蛋白纯化相关产品
| 货号 | 名称 | 规格 |
| BR100530 | Series S Sensor Chip CM5 | 3片装 |
| 29149603 | Series S Sensor Chip CM5 | 10片装 |
| BR100050 | Amine Coupling Kit | / |
PART2 FGL1在体外和体内通过LAG3抑制抗原介导的T细胞反应
LAG3在静止的T细胞上不表达,但是可以在多种抗原刺激的情况下被上调。通过流式分析发现,FGL1融合蛋白不能结合静止的T细胞,但是可以和野生型小鼠的激活的T细胞结合,而不能结合LAG3-KO的小鼠的激活的T细胞(Fig. 2a)。在anti-CD3刺激的情况下,FGL1可以部分抑制野生型脾脏T细胞的增殖,而这个抑制在LAG3-KO的脾脏T细胞中被减弱,表明FGL1的抑制作用依赖于LAG3(Fig. 2b)。
同时,FGL1剂量依赖性的抑制LAG3过表达的3A9 T细胞的IL-2的抗原特异性诱导,相比于低内源性LAG3表达的亲代3A9 T细胞(Fig. 2c)。作者构建了一个针对于FGL1的中和抗体(177R4),可以阻滞FGL1与LAG3过表达的T细胞结合。177R4和C9B7W都能阻止3A9-LAG3细胞中FGL1对IL-2诱导的抑制(Fig. 2d)。CD8+ OT-1转基因的T细胞转染至同源的小鼠中,使用鸡的卵清蛋白多肽抗原免疫,通过检测到血浆水平TNF-α and IFN-γ的上调表明,FGL1中和抗体177R4显著促进抗原特异性的OT-1 T细胞的激活,效果等同于LAG3中和抗体(Fig. S2c-d)。 
Fig.2 a-d FGL1调节LAG3依赖的T细胞抑制
△点击放大图片

Fig.S2 c-d FGL1中和抗体增强体外的抗原特异性T细胞反应
△点击放大图片
综上表明,FGL1是一个LAG3的抑制性配基。
PART 3 Fgl1缺陷的小鼠缓慢的发展自发的自身免疫系统症状
作者构建了一个FGL1-KO的小鼠系。在野生型小鼠中,FGL1 mRNA在肝脏中被检测出而不是在其他组织或者造血细胞中。可溶的FGL1在小鼠血液中被检测出,而不能在FGL1-KO的小鼠的血浆或者肝脏中被检测出来(Fig. S3b-c)。
FGL1-KO的小鼠具有正常的外观,器官大小和繁殖,表明FGL1不能总体的影响小鼠生长和发育。然而40%的FGL1-KO小鼠在8个月之后会出现自发性皮炎,显示出皮肤中的淋巴细胞浸润(Fig. S3d-e)。在14-16个月,5/8的雌性FGL1-KO小鼠,没有雄性小鼠,在血浆中抗双链DNA自身抗体的水平增加,相比于野生型小鼠(Fig. S3f)。这些发现和FGL1是一个免疫抑制性分子的功能相一致。
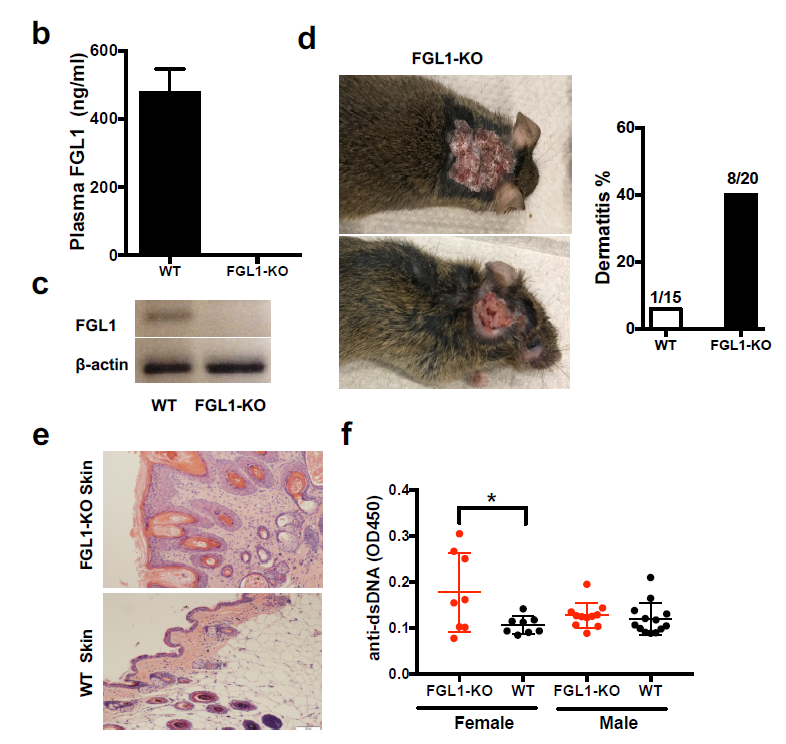
Fig.S3 b-f FGL1-KO小鼠自身免疫表现
△点击放大图片
为了评估KO小鼠中免疫系统的总体变化,作者用mass cytometry(CyTOF)描绘了小鼠的外周血细胞。作者检测了全部的CD45+造血细胞,其中central memory-like CD8+ T细胞亚群(cluster 14–15, CD44+ CD62L+ Ly6C+ CD127med Tbet+ Eomes+)有显著的增加,而两个B细胞亚群(cluster 2 and 4)下降(Fig. 3a-e)。
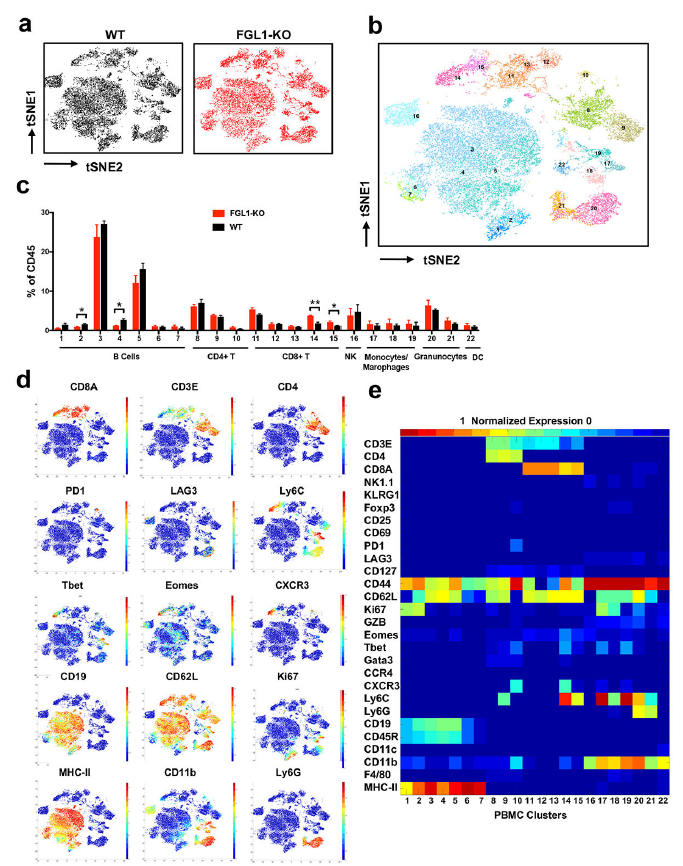
Fig.3 a-e FGL1-KO小鼠免疫细胞表型
△点击放大图片
这些结果表明内源的FGL1不影响小鼠的发育和生长,而可能参与自身免疫的调节和免疫稳态的调节。
PART 4 在小鼠模型中Fgl1的沉默促进T细胞免疫对抗肿瘤生长
FGL1-KO和LAG3-KO小鼠皮下接种同系的小鼠MC38结肠癌细胞。和LAG3-KO的小鼠一样,FGL1-KO的小鼠和正常小鼠相比肿瘤生长速度明显减慢(Fig. 4a)。而FGL1和LAG3中和抗体也显著减慢了MC38小鼠结肠细胞和Hepa1-6小鼠肝脏细胞的生长(fig. 4c)。相反,FGL1和LAG3中和抗体的抗肿瘤效应在Rag1-KO C57BL/6小鼠(缺乏T细胞和B细胞)中无效(Fig. 4d)。进一步验证发现,在MC38肿瘤模型中,用特定的抗体中和CD8+ 或者 CD4+ T细胞,可以完全的消除FGL1和LAG3中和抗体的抗肿瘤效应,表明这些中和抗体的抗肿瘤效应依赖于CD8+和CD4+ T细胞。
为了排除LAG3其他的配基与FGL1作用影响LAG3中和抗体的抗肿瘤效应,作者在FGL1-KO的小鼠中验证LAG3中和抗体。在野生型小鼠中,LAG3中和抗体抑制了MC38肿瘤生长,而这个抗肿瘤的效应再FGL1-KO的小鼠中被消除(Fig. 4e)。而这种效应也依赖于LAG3,因为FGL1中和抗体在LAG3-KO小鼠中对肿瘤生长也没有额外的影响(Fig. 4f)。因此,FGL1中和抗体的抗肿瘤效应依赖于LAG3,而LAG3中和抗体的效应依赖于FGL1而不依赖于MHC-II或者LAG3的其他配基。这些结论表明FGL1是一个LAG3的主要配基来诱导T细胞抑制作用和免疫逃逸。

Fig.4 a-f 在小鼠模型中FGL1/LAG3相互作用的消除抑制肿瘤生长
△点击放大图片
流式实验分析表明,在FGL1-KO的肿瘤中,CD45+白细胞数量明显增加,其中CD44+ CD62L- PD-1+ Gata3+ effector memory-like CD4+ TIL和CD44+ Ly6C+ memory-like CD8+ TIL数量明显增加(Fig. 5b-e)。相反,Treg NK或者B细胞没有明显改变(Fig. 5b-e)。

Fig.5 a-e 在FGL1-KO小鼠中肿瘤浸润T细胞的增殖
△点击放大图片
综上表明,通过转基因敲除或者中和抗体使FGL1/LAG3沉默会促进肿瘤免疫,通过在肿瘤微环境中刺激T细胞扩增和优先激活。
PART 5 高血浆FGL1和病人在anti-PD治疗中效果更差有关
为了检测FGL1抑制肿瘤免疫是否不依赖于B7-H1/PD-1通路,作者检测了在转移性NSCLC病人中,血浆基准FGL1水平和B7H1/PD-1阻滞治疗(anti-PD)中的关联。作者发现,更高的血浆FGL1水平和NSCLC病人anti-PD治疗效果更差有关(Fig. 6e)。结果表明,FGL1/LAG3相互作用独立于B7-H1/PD-1通路,潜在的引起在人癌症中anti-PD治疗抵抗。
作者进一步使用了MC38肿瘤模型检测在B7-H1/PD-1通路阻滞存在的情况下anti-FGL1/anti-LAG3的作用。对于第6天接种MC38肿瘤的小鼠使用中和抗体进行处理。如果单独使用,anti-FGL1,anti-LAG3和anti-B7-H1抗体会减缓肿瘤生长和少量延长存活率(Fig. 6g)。然而,anti-FGL1或者anti-LAG3抗体和anti-B7-H1抗体联用会显著的提高存活率和降低肿瘤负荷,和使用单个抗体治疗相比(fig. 6g-h)。

Fig.6 d-h 在人癌症中FGL1的上调和不良预后有关
△点击放大图片
综上表明,FGL1/LAG3通路是一个独立的肿瘤免疫逃逸机制,阻断其相互作用和anti-PD疗法具有协同作用。
其他相关产品
| 货号 | 产品名称 |
| 558739 | Ms CD44 Pure KM114 100ug |
| 550426 | Ms CD127 NALE SB/14 500ug |
| 550609 | Ki-67 IHC Pure B56 1mL |
| 553295 | Ms CD28 Pure 37.51 500ug |
| 553864 | Ms CD25 NALE PC61 500ug |
| 553783 | Ms CD19 Pure 1D3 500ug |
| 565409 | Ms F4/80 Pure T45-2342 100ug |
| 550280 | Ms CD4 IHC Pure RM4-5 1mL |
| 550286 | Ms CD45R IHC Pure RA3-6B2 1mL |
| 553799 | Ms CD11c Pure HL3 500ug |
| 610197 | CD95/Fas/APO-1 Pure 13/Fas 50ug |
| 130-093-235 | GentleMACS组织处理器 |
| 550749 | Hu Th1/Th2 CBA Cytokine Kit |
| 29122S | PD-L1 (405.9A11) Mouse mAb |
| 553294 | Ms CD28 NALE 37.51 500ug |
| 553057 | Ms CD3e NALE 145-2C11 500ug |