


视频素材来源于LabTube https://www.labtube.tv/video/MTA2MTMz
版权归原作者所有
自噬中的信号通路
- 1
- 2
- 3
-
自噬中的信号通路介绍

△点击放大图片
巨自噬,通常也称作自噬,是一种催化过程,导致细胞浆主要内容物的自噬性溶酶体降解、异常蛋白聚集和细胞器过剩或受损。营养不足的状态通常会激活自噬,但自噬也与许多生理和病理过程有关,比如发育、分化、神经退行性疾病、应激、感染以及癌症。mTOR 激酶是诱导自噬的重要调节分子,激活的 mTOR(Akt 和 MAPK 信号转导)可抑制自噬,而 mTOR(AMPK 和 p53 信号转导)的负性调节则促进自噬。三种相关的丝氨酸/苏氨酸激酶,UNC-51-激酶-1、-2 和 -3 (ULK1、ULK2、UKL3),与酵母 Atg1 具有同样的作用,可作为 mTOR 复合体的下游。ULK1、ULK2 与一种自噬相关 (Atg) 基因产物 (mAtg13) 的哺乳动物同源体,以及骨架蛋白FIP200 与酵母 Atg17 直接同源)共同形成一个较大的复合体。III 级 PI3K 复合体,包含 hVps34、 Beclin-1(酵母 Atg6的一种哺乳动物同源体)、 p150(酵母 Vps15的一种哺乳动物同源体)以及 Atg14 样蛋白 (Atg14L 或 Barkor) 或抗紫外辐射相关基因(UVRAG),为诱导自噬所必须。 线粒体自噬是一种选择性自噬过程,可特异性地从细胞移除受损或不需要的线粒体部分。PINK 蛋白在健康状态下通过 PARL 的作用持续降解,而在线粒体损伤时,PINK 稳定并招募 E3 连接酶 Parkin ,以启动自噬。
线粒体膜蛋白通过 Parkin 的多聚泛素化作用导致自噬连接蛋白 SQSTM1/p62、NBR1 和 Ambra1 聚集,并通过 LC3-作用区域 (LIR) 结合到 LC3上。另外,BNIP3 和 BNIP3L/NIX 也含有 LIRs,可通过非泛素化机制直接聚集自噬机制因素,从而在某些细胞类型中诱导自噬体的形成。 -
mTOR是细胞自噬的核心

△点击放大图片
mTOR信号通路是调控细胞生长与增殖的一个关键通路, 该通路将营养、能量状态及生长因子信号整合在一起, 并作为主要的调控因子参与细胞自噬过程。mTOR有两种不同的复合体, 即mTORC1和mTORC2。
mTORC1 通过磷酸化使形成的自噬调节复合物 (由 ULK1 和其互作蛋白 Atg13、FIP200、Atg101 等形成) 失活,从而影响自噬小体的生物发生。在营养丰富的条件下,mTORC1 通过介导 ULK1 (Ser637 和 Ser757) 和 Atg13 (Ser258) 特定位点磷酸化,抑制 ULK1 复合物的自噬促进激酶 (Autophagy-promoting kinase) 活性。
在饥饿和细胞应激期间,mTORC1 活性被抑制,于是与 ULK1 分离。因此,ULK1 和 Atg13 的特定位点的磷酸化被解除。同时,ULK1 复合物通过 Thr180 处自磷酸化而变得活跃,并磷酸化 Atg13、FIP200、Atg101 和其他 Atg 蛋白。活跃的 ULK1 复合物随后转移到内质网的隔离膜上,自噬启动。 -
细胞自噬中的AMPK通路与P53
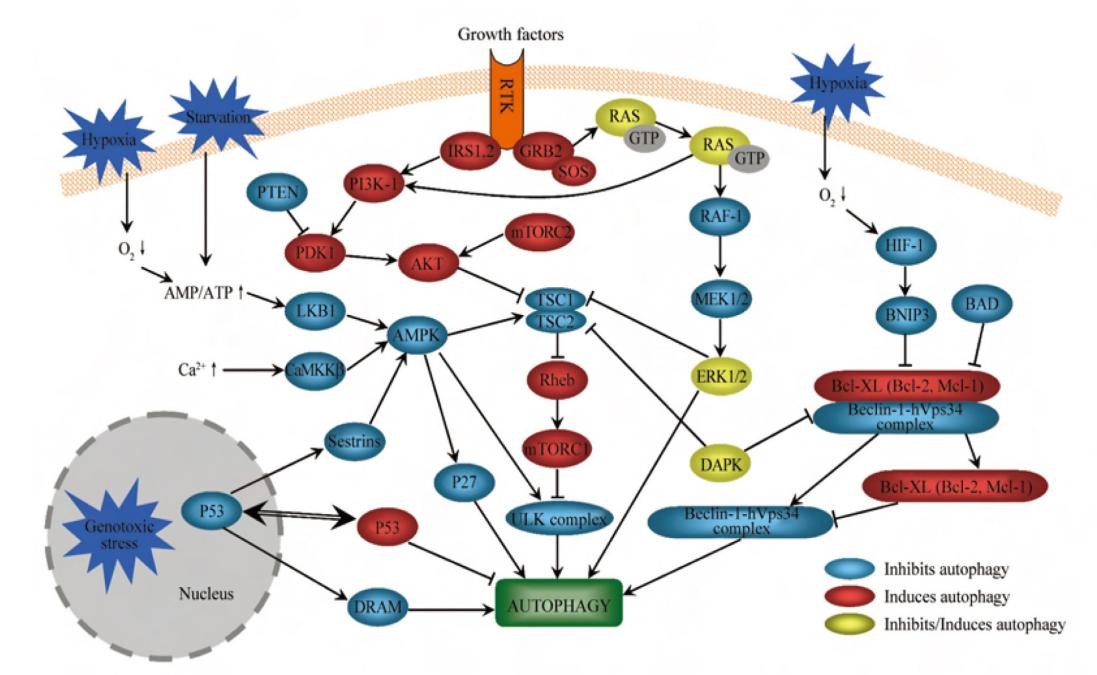
△点击放大图片
苷酸活化蛋白激酶(AMP-activated protein kinase, AMPK)是能量代谢变化的感受器, 在能量代谢过程中起重要的调节作用。在营养缺乏的情况下, ATP/AMP的比例下降引起LKB1的激活, 进而激活AMPK。与AKT通路的作用相反, AMPK可以磷酸化激活TSC1/2复合体, 进而抑制mTORC1的活性。
AMPK还可以通过直接磷酸化Raptor, 使Raptor脱离mTORC1复合体,进而抑制mTORC1的活性。AMPK是自噬的重要正调控因子, 在压力存在的情况下, LKB1-AMPK通路磷酸化细胞周期抑制剂p27kip1来激活自噬反应。细胞内游离Ca2+的浓度增加可以通过激活Ca2+/CaMKKβ和TAK1通路, 进而激活AMPK, 最终通过抑制mTOR诱导自噬的发生。最新的研究表明, 营养缺乏会促进AMPK与ULK1的PS结构域结合, 使ULK1的Ser/Thr丰富 区多位点磷酸化, 从而激活ULK1, 直接调节细胞自噬。
p53是一种重要的抑癌基因, 对自噬的调控作用是双重的。DNA损伤促进p53表达, p53通过激活MPK通路, 诱导了自噬的发生。
p53入核后会增加DRAM (damage-regulated autophagy modulator)的转录进而促进自噬, 胞浆内p53的减少也提高了自噬的水平。